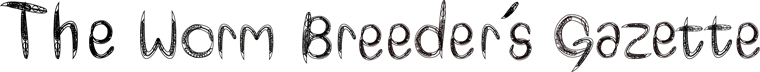
In C. elegans dwelling and roaming activities are two examples of distinctive behaviors. During the dwelling phase worms have an intermittent backward and forward movement and present a low/high speed of locomotion. However throughout the roaming phase worms have predominantly high speed forward movement with few changes of direction (Fujiwara et al., 2002; Shtonda and Avery, 2006). These two different behaviors are controlled by the presence or absence of food in the environment (Fujiwara et al., 2002). Both states seem to be regulated in a different way, thus while dwelling is induced by internal metabolic perceptions, roaming is mediated by stimuli detected via the amphid neurons (Ben Arous et al., 2009).
Fig. 1 shows that the roaming behavior of nrx-1-deficient mutants was almost annulled when worms were transferred from OP50 bacteria plates to plate without food. This contrasts with the behavior of N2 wild type strain. Interestingly, transgenic expression in the nrx-1 deficient mutant of cDNAs coding human alpha-NRXN-1 or beta-NRXN-1, all driven by the C. elegans nrx-1 promoter, rescued the mutant phenotype of the nematode (Fig. 1). In these experiments nrx-1(tm1961) transgenic animal expressing beta-NRX-1 from the nematode were used as control. To quantify the exploratory capacity of the worms, twenty L4 larval stage animals of each genotype grown on OP50 E. coli in fresh NGM plates were picked and placed in the middle of a 9 cm plate containing 8 ml of agar medium (2% agar, 0.25% v/v Tween 20, 10 mM HEPES pH 7.2) without bacteria. Plates were incubated from 5 to 7 days at 20 ºC. The fingerprints of the worms were recognized by sight as a consequence of their roaming, and they were quantified by scanning the surface of agar dishes at 600-ppp resolution and analyzing the images in “Image J” software using custom scripts. Images were turned into binary format converting them through a predetermined threshold, and finally a particles analysis was performed. Data was normalized with respect N2 wild-type strain.
A previous report had shown that human and rat neuroligin-1 were functional in C. elegans (Calahorro and Ruiz-Rubio, 2012). Together, these results suggest that the functional mechanism underpinning both neuroligin and neurexin in the nematode are comparable to human. In this sense the nematode might constitute a simple in vivo model for understanding basic mechanisms involved in some neurological diseases.
Figures
References
Ben Arous J, Laffont S, Chatenay D. (2009) Molecular and sensory basis of a food related two-state behavior in C. elegans. PLoS One, 4, e7584. 
Calahorro F, Ruiz-Rubio M. (2012) Functional phenotypic rescue of Caenorhabditis elegans neuroligin-deficient mutants by the human and rat NLGN1 genes. PLoS One, 7, e39277.
Fujiwara M, Sengupta P, McIntire SL. (2002) Regulation of body size and behavioral state of C. elegans by sensory perception and the EGL-4 cGMP-dependent protein kinase. Neuron, 36, 1091-1102.
Shtonda BB, Avery L. (2006) Dietary choice behavior in Caenorhabditis elegans. J Exp Biol, 209, 89-102.