The ease of generating transgenic animals is one of the key experimental advantages of C. elegans as a model organism. Typically, microinjection of DNA encoding the desired transgene results in heritable extra-chromosomal arrays that provide robust expression in the somatic tissues of C. elegans. Achieving expression in the germline with such arrays is however, less successful due to silencing of repetitive sequences in this tissue. In order to “dilute” the repetitiveness of the array to achieve germline expression, addition of N2 genomic or foreign DNA in the array has been shown to be effective (Kelly et al. (1995) WBG 14(1): 64; Kelly et al., 1997; Lam et al. 2006). Another way to achieve germline expression is the production of low copy-number arrays using methods such as the generation of single-copy arrays by microparticle bombardment or transposon-mediated chromosomal integration approaches (Praitis et al., 2001; Frokjaer-Jensen et al., 2008). However, in contrast to microinjection protocols, these approaches often produce low expression levels of reporters such as fluorescent proteins and they also require the use of specific genetic backgrounds.
In our lab we have been able to obtain stable germline expression from several extrachromosomal and integrated transgenic reporters using a microinjection-based protocol, in which we include either sonicated or PvuII digested E. coli genomic DNA as carrier DNA in the injection mix.
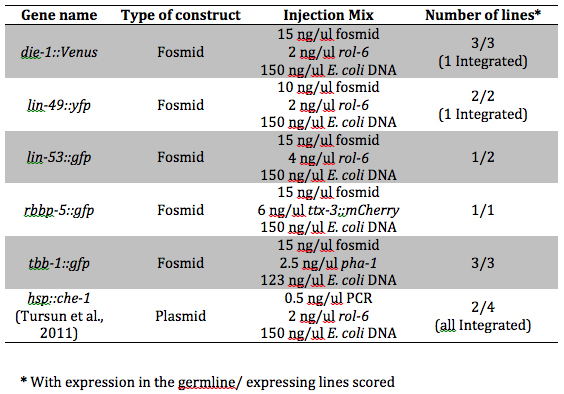
We obtain E. coli genomic DNA using standard methods or commonly used genomic DNA kits. We sonicate the DNA to obtain a smear of DNA sizes centered at around 1kb. Fragmentation of E. coli genomic DNA can be also performed by complete PvuII digestion. Both ways give stable lines with germline expression in our hands. We then pass the fragmented genomic DNA through DNA purification columns. The final concentration of bacterial DNA in the injection mix is from 100-150 ng/ul and reporters and co-injection markers are linearized and injected at the concentrations shown in Table 1. We have successfully obtained germline expression of small reporters (plasmid based) as well as fosmid reporters. We also found no effect on germline expression using either DNA methylation competent or deficient E. coli strains as genomic DNA source.
In Table 1, we show transgenes injected using our protocol that gave stable germline expression, some of which had previously been characterized with respect to copy number using aCGH (Cochella et al. (2011) WBG 18(4)). In this table there are transgenes that have low copy number, like tbb-1 (otEx4548) (mean copy number 5/array), and high copy number, like hs::che-1 (otIs284) (mean copy number 11/chromosome). This suggests that for the protocol that we follow, copy numbers within the range that we have obtained can support germline expression.
Although the addition of bacterial DNA to arrays is not novel, as far as we have followed these strains, we do not observe silencing of reporters after several generations as noted before (Kelly et al. (1995) WBG 14(1): 64). In addition, using bacterial DNA instead of N2 genomic has also the advantage of not introducing wild-type worm genes into mutant backgrounds.
We cannot exclude that expression of some genes may be more sensitive to copy number and/or genomic context and therefore may still require low/single-copy insertions to produce expression in the germline. However, we believe that this is a robust and simple enough approach to be considered as the first thing to try when germline expression is expected/required. Therefore, we hope that our method will help C. elegans researchers to obtain germline expression of transgenes using microinjection techniques.
Figures
References
Frokjaer-Jensen C, Davis MW, Hopkins CE, Newman BJ, Thummel JM, Olesen SP, Grunnet M, Jorgensen EM. (2008) Single-copy insertion of transgenes in Caenorhabditis elegans. Nature Genet 40, 1375-1383. 
Kelly WG, Xu S, Montgomery MK, Fire A. (1997) Distinct requirements for somatic and germline expression of a generally expressed Caernorhabditis elegans gene. Genetics 146, 227-238.
Lam N, Chesney MA, Kimble J. (2006) Wnt signaling and CEH-22/tinman/Nkx2.5 specify a stem cell niche in C. elegans. Curr Biol 16, 287-295.
Praitis V, Casey E, Collar D, Austin J. (2001) Creation of low-copy integrated transgenic lines in Caenorhabditis elegans. Genetics 157, 1217-1226.
Tursun B, Patel T, Kratsios P, Hobert O. (2011) Direct conversion of C. elegans germ cells into specific neuron types. Science 331, 304-308.