Embryo series courtesy of Einhard Schierenberg
Embryo series courtesy of Einhard SchierenbergAbstract
The mechanism of action of volatile anesthetics remains an enigma, despite their worldwide use. The nematode C. elegans has served as an excellent model to unravel this mystery. Genes and gene sets that control the behavior of the animal in volatile anesthetics have been identified, using multiple endpoints to mimic the phenomenon of anesthesia in man. Some of these studies have clear translational implications in more complicated organisms.
The first successful public demonstration of reversible loss of consciousness by a volatile anesthetic occurred on October 16, 1846. Administration of diethyl ether allowed the removal of a neck tumor from a quiescent and pain free patient. Not only did this stunning demonstration revolutionize medical practice by changing the scope and frequency of surgery, but it was viewed as a triumph over pain and hailed as a gift to humanity. Oliver Wendell Holmes, who went on to become the dean of the Harvard School of Medicine, coined the phrase “anesthesia” in order to give a name to something that had never been conceived as possible by physicians prior to that time (Fenster, 2001). The hallmarks of volatile agents as complete general anesthetics include amnesia, analgesia, muscle relaxation, and loss of consciousness, which are not provided in toto by any other single class of medicinal agents (Zhang et al., 2000). In the past 150 years a number of seemingly unrelated volatile compounds have been used as volatile anesthetics. From heavily substituted halogenated alkanes and ethers to elemental noble gases, volatile anesthetics are still a mainstay of modern medicine (Zhang et al., 2000). However, their volatility, lack of physical similarity to each other, and apparent weak affinity for their target(s), has hampered understanding of their mechanism(s) of action at the molecular level. Although empirically well understood in their clinical administration, volatile anesthetics do not yet lend themselves well to rational drug design. Each volatile anesthetic currently in use is associated with its own characteristic side effects and limitations.
Early work on the mechanism of action of volatile anesthetics took note of the high lipid solubility of this class of compounds, as well as the high amount of lipid staining in the brain. By the turn of the 20th century, Meyer and Overton independently noted that volatile anesthetics increase in potency as a function of their solubility in olive oil, a heterogeneous collection of non-polar lipids. This held true for all species in which anesthetics were tested, over four to five orders of magnitude of lipid solubility (Meyer, 1899; Overton, 1901; Tanifuji et al., 1977). Termed the Meyer-Overton rule, the correlation led to early hypotheses about how volatile anesthetics work. A plot of this relationship could be used to derive an interesting corollary. Assuming that the anesthetic target and/or its milieu shares physical properties with the solvent olive oil, all inhaled anesthetics, are present in equimolar concentrations at the anesthetic site when they have reached their effective volatile concentrations (Tanifuji et al., 1977). It was therefore viewed that lipid solubility was the parameter that controlled access to the anesthetic target, but that once actually reaching their site of action, all inhaled anesthetics were equally potent. Also, volatile anesthetics are generally additive in their effects, i.e. a mixture of a half dose of two different volatile anesthetics was in fact equal to a full dose of either drug in isolation. In this manner a unitary theory of narcosis was born, in which all volatile anesthetics work at the same single target in all species (Deady et al., 1981). Most theories originally favored a fairly non-specific insinuation of the volatile anesthetic into the lipid bilayer of excitable cell membranes, causing inhibition of neuronal conduction. Changes in membrane density (critical volume hypothesis) and membrane fluidity were invoked as the mechanism of action of volatile anesthetics (Suezaki et al., 1990). Although these physical changes definitely occur in membranes in the presence of volatile anesthetics, relatively small increases in temperature can mimic these effects on membrane fluidity and density without causing anesthesia. For this and other reasons (see below), these early models that hypothesized a general, non-specific interaction of volatile anesthetics with cell membranes fell into disfavor.
Through the work of Franks and Lieb and others, it was shown that in fact a Meyer-Overton plot could be obtained by substituting octanol for olive oil as the solvent in which volatile anesthetic solubility was measured (Abraham et al., 1991). An equally good fit exists between potency and solubility of volatile anesthetics in this polar molecule. In addition, volatile anesthetics can inhibit luciferase in a cell free extract; their potency of inhibition correlates well with their potency as anesthetics (Moss et al., 1991). Proteins are currently favored as potential targets of volatile anesthetics, and many molecules have been proposed as anesthetic targets. In particular, various membrane ion channels and receptors are hypothesized to be the site of action of volatile anesthetics. Volatile anesthetics affect the functions of multiple membrane proteins, most notably ligand gated ion channels (GABAA receptors, glutamate receptors), voltage gated ion channels, G-protein coupled receptors and various other ion channels (reviewed in Evers and Crowder, 2006). Currently no single protein or channel has emerged as the likely anesthetic target to the exclusion of all others. Current debate entertains the possibility that multiple targets may exist for volatile anesthetics. The extremely steep dose-response curves seen for this class of molecules are certainly most consistent with multiple sites of action. If this is the case, then the relative importance of each potential target cannot be easily determined using an in vitro approach.
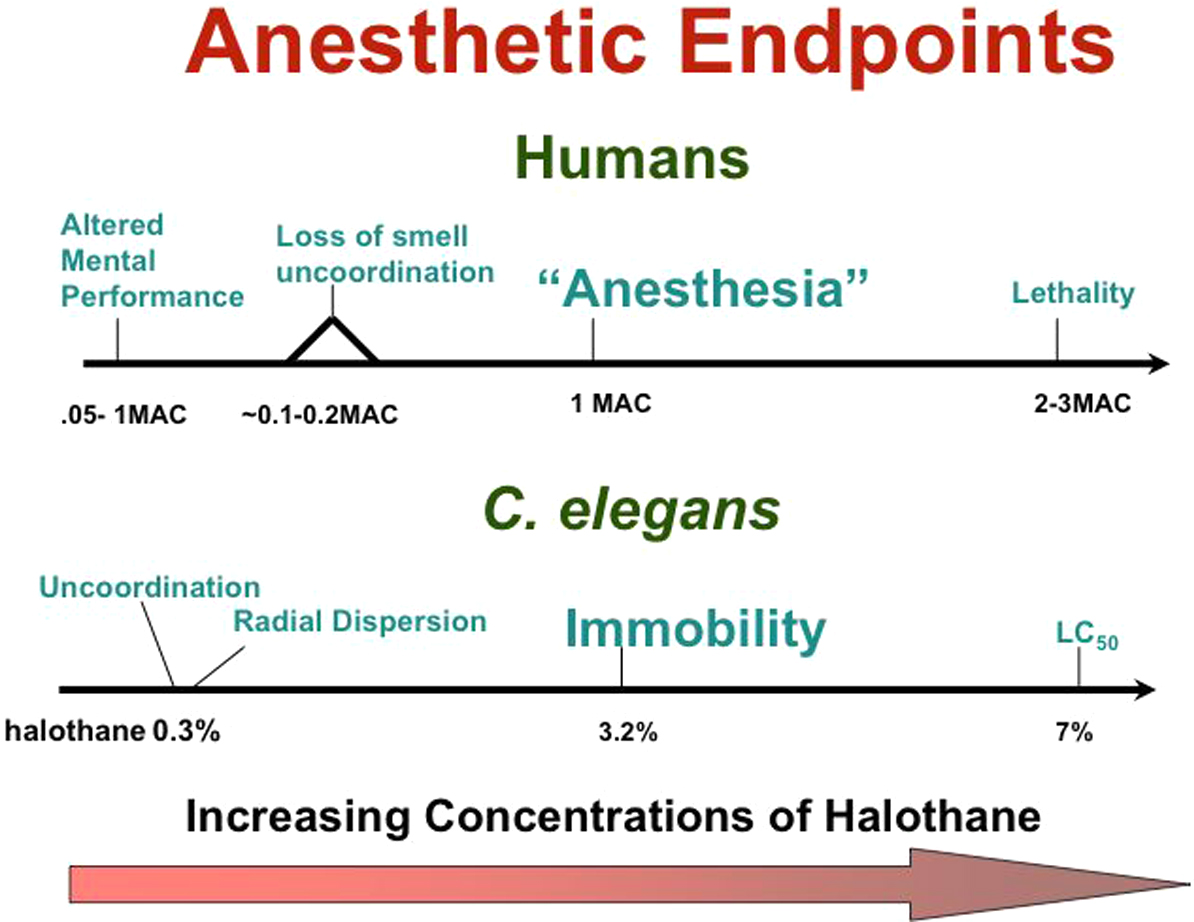
“Anesthesia” is clearly a whole animal phenomenon. Anesthetics as volatile, lipid soluble chemicals can perturb virtually any experimental system into which they are introduced. Therefore a model organism such as the nematode is invaluable in understanding how the volatile anesthetics work. C. elegans has multiple behaviors which can be used as anesthetic endpoints (Morgan and Cascorbi, 1985; Crowder et al., 1996). When exposed to volatile anesthetics, wild-type nematodes first increase movement, become ‘excited’, and lose their response to volatile attractants. This behavior proceeds to a progressive lack of coordination, followed by immobility and unresponsiveness to a tap to the snout (see Movie 1; Morgan and Cascorbi, 1985). This progression of neurologic changes is reminiscent of the neurological responses seen in patients, who experience an excitatory phase upon induction of general anesthesia. Excitation gradually becomes progressively uncoordinated movement that eventually slows to complete immobility in response to a surgical stimulus. Several different endpoints have been used to measure anesthetic effects in C. elegans. The two most commonly used have been (1) some form of uncoordination or change in motion in response to anesthetic exposure and (2) immobility. Both endpoints can demonstrate a Meyer-Overton relationship, i.e. potency strictly follows lipid solubility; in each assay then the order of potency of gases is very similar to that seen in humans. The mechanisms of the two anesthetic actions are distinct and likely correspond to different components of the set of complex behavioral effects seen in the vertebrate nervous system (see Figure 1). Insight concerning the mechanisms of action of volatile anesthetics will probably come from studies of several endpoints.
|
Figure 1. Absolute and relative doses required to achieve several endpoints in humans and in C. elegans. The concentrations of anesthetic necessary to cause immobility are higher in the nematode than in humans. Note, also, the relative positions of anesthetic endpoints in humans compared to the relative positions for endpoints in the nematode. It is not known which endpoints in the nematode correspond to specific endpoints in humans.
Concentrations of volatile anesthetics that produce immobility in response to a surgical stimulus in humans do not immobilize worms. However, at these concentrations worms do exhibit an easily scored sluggishness and uncoordination (Crowder, 1996). Using radial dispersion as an endpoint, Crowder and colleagues (Crowder et al. 1996; van Swinderen et al., 1999; van Swinderen et al., 2002) screened nematodes carrying previously identified mutations in neuronal proteins of C. elegans for altered sensitivity to anesthetics. They found that mutations in syntaxin (unc-64) caused either resistance or hypersensitivity to volatile anesthetics. Syntaxin is a member of the protein complex that controls presynaptic vesicular fusion with the cell membrane, resulting in neurotransmitter release into the synaptic cleft (Koushika and Nonet, 2000). This complex, the SNARE complex, includes the syntaxin-binding proteins synaptobrevin and SNAP-25 (Nonet et al., 1998; Rand and Nonet, 1997). The syntaxin allelic variation was striking, particularly for isoflurane, where a 33-fold range of sensitivities was seen. Mutations causing hypersensitivity reduced neurotransmitter release, while the resistant class generally increased release. However, one mutation dominantly conferred resistance to the volatile anesthetics, isoflurane and halothane, but decreased neurotransmitter release. The authors concluded that syntaxin is a possible target of volatile anesthetics, and that anesthetics ordinarily work by decreasing neurotransmitter release via syntaxin binding. Later data indicated that mammalian syntaxin is capable of binding halothane (Nagele et al., 2005a).
Using a different set of endpoints (velocity of locomotion and the body bend assay), Crowder and Nonet tested anesthetic sensitivity of mutants isolated in a screen for enhancers of transmitter release (Wang et al., 2001; Hawasli et al., 2004). They screened for suppression of a hypomorphic unc-64 to facilitate identification of mutations that enhanced syntaxin function. The largest group of genes included loss of function alleles of slo-1, which encodes the calcium activated potassium channel (BK channel); not surprisingly, slo-1 animals were resistant to halothane (see Table 1). Wang showed that this channel negatively regulates neurotransmitter release in C. elegans (Wang et al., 2001). slo-1 alleles have also been identified in screens for resistance to ethanol (Davies et al., 2003) which has been thought to share some mechanistic similarities to the volatile anesthetics. Hawasli et al. (2004) also found that mutations in unc-43 suppressed the syntaxin mutants (unc-43 is a calcium/calmodulin dependent protein kinase). Mutations in unc-43 did not change volatile anesthetic sensitivity in isolation (see Table 1).
Table 1. Mutations with multiple interactions that affect anesthetic sensitivity in C. elegans.
| Mutants | Product | Nonanesthetic phenotype | Endpoint | Anesthetic phenotype | Interactions | Comments |
|---|---|---|---|---|---|---|
| unc-64 | Syntaxin | sluggish | Radial dispersion | Allele specific changes | goa-1,slo-1, unc-43,egl-30 | Postulated direct target |
| slo-1(lf) | BK channel | Velocity of locomotion | Resistant | Suppress unc-64 | ||
| goa-1(lf) | Alpha subunit of Go | hyperactive | Radial dispersion | Resistant | ||
| egl-30(gf) | Alpha subunit of Gq | Velocity of locomotion | egl-30(lf) hyper-sensitive | Suppress unc-64 | ||
| unc-43 | Calcium dependent protein kinase | Velocity of locomotion | None | Suppress unc-64 | ||
| unc-79 | Novel | fainter | immobility | Hyper-sensitive to specific anesthetics | unc-1, unc-8, unc-7, unc-9, unc-24 | |
| unc-80 | Unknown | fainter | immobility | Hyper-sensitive to specific anesthetics | unc-1, unc-8, unc-7, unc-9, unc-24 | |
| unc-1(lf), unc-24(lf) | Stomatins | kinker | immobility | Hyper-sensitive to ether | Suppress fainters | gf alleles of unc-1 hyper-sensitive to all VAs |
| unc-7(lf) unc-9(lf) | Gap junction proteins | kinker | immobility | Hyper-sensitive to ether | Suppress fainters | |
| unc-8(gf) | Degenerin/ENaC | kinker | immobility | Hyper-sensitive to ether | Suppress fainters | lf alleles suppress unc-79 |
| nca-2,nca-1 | Cation channels | nca-2;nca-1 is a fainter | immobility | Hyper-sensitive to specific anesthetics | unc-1, unc-8, unc-7, unc-9, unc-24 | Protein product lost in unc-79, unc-80 |
| Note: Mitochondrial mutations are not listed. See text. | ||||||
Finally, and perhaps most remarkably, a gain-of-function mutation in egl-30 suppressed the hypersensitive syntaxin mutants, while loss-of-function mutants in egl-30 increased anesthetic sensitivity (Hawasli et al., 2004). egl-30 encodes the a subunit of the G protein Gq, previously shown to positively regulate locomotion and other elements of behavior (Miller et al., 1999; Lackner et al., 1999). These results are particularly interesting in that the same laboratory earlier showed that loss of function mutations in goa-1 led to volatile anesthetic resistance (van Swinderen et al., 2001). The results are consistent with goa-1 and egl-30 having antagonistic functions in regulating neurotransmitter release and volatile anesthetic sensitivity of this endpoint. In summary, each of these mutations indicates that the general activity of the presynaptic region is of importance in determining anesthetic sensitivity. Neurotransmitter release may be elevated or depressed, with resulting changes in anesthetic sensitivity. The authors also concluded that SLO-1 and EGL-30 were unlikely to encode a volatile anesthetic target.
Corroborating their work in C. elegans, Crowder and colleagues later showed that isoflurane inhibited the presynaptic action potential in the rat brainstem at the Calyx of Held (Wu et al., 2004). This inhibition decreased glutamate release and, in turn, the excitatory postsynaptic potential as a secondary effect. The authors concluded that the presynaptic effects contributed to the inhibition of postsynaptic currents. The same laboratory expressed rat orthologues of many of the SNARE proteins in a heterologous system, and found that both the SNARE complex and the syntaxin protein were capable of binding both halothane and isoflurane (Nagele et al., 2005a).
In closely related work, Crowder and colleagues studied the action of nitrous oxide (laughing gas) in C. elegans (Nagele et al., 2004). They found that nitrous oxide produced a behavior similar to that produced by loss-of-function mutations affecting either the AMPA or NMDA glutamate receptors in the worm. Only the nmr-1 mutant animal (the nmr-1 gene codes for the NMDA receptor) was resistant to the effects of nitrous oxide. They concluded that the NMDA receptor was required for the effects of nitrous oxide. The same laboratory also studied the effects of the volatile anesthetic xenon and found that the non-NMDA glutamate receptor glr-1 was necessary for the anesthetic effects of the drug (Nagele et al., 2005b).
Interestingly, and in contrast to studies in syntaxin by this group, these last two studies implicate postsynaptic activity as key to controlling sensitivity to xenon and nitrous oxide. Thus, it appears that different volatile anesthetics may function differently in causing uncoordination and decreased speed of locomotion.
In human patients, the effective concentration of a volatile anesthetic necessary for surgical anesthesia is termed “MAC” for “minimal alveolar concentration.” It is the concentration of a gas that, in 50% of patients, will cause a patient to remain immobile during a surgical stimulus (Regan and Eger, 1967). In an effort to mimic MAC in worms, Morgan and Sedensky used reversible immobility as an endpoint to study control of anesthetic behavior in C. elegans (Morgan and Cascorbi, 1985). This screen uses higher concentrations of anesthetics than those safely used in patients, although the relative order of their potency is nearly identical to that seen in humans. In this assay, the Meyer-Overton relationship holds true for a range of lipid solubilities that varies by nearly 4 orders of magnitude. Mutations affecting immobility in anesthetics identified an interacting set of genes that have been ordered relative to each other, and has yielded some surprising correlates in studies in other species. However, it is important to note that the gene products may not directly interact with each other or be in a common metabolic pathway. Apparently epistatic mutations may actually affect cells or functions which are themselves “downstream” in causing immobility. In addition, those genes identified in screens for uncoordination caused by anesthetics generally have little effect in immobility assays; vice versa - genes identified in screens for immobility have not been found in screens using radial dispersion as an endpoint.
Screening for hypersensitivity to the anesthetic halothane, Sedensky and Morgan identified a new allele of unc-79(III) which caused the animal to be very hypersensitive to a number of gases (Morgan and Cascorbi, 1985; Sedensky and Meneely, 1987). In general, using immobility as an endpoint gives steep dose-response curves, such that unc-79 animals are absolutely immobile in 2% halothane, while N2 animals move vigorously. However, an unexpected feature of unc-79 animals was that they did not respond equally to all volatile anesthetics. They are hypersensitive to halothane (and the most lipid soluble volatile anesthetics), but react like wildtype to a second set of volatile anesthetics, and are resistant to yet other gases. This ability to separate out anesthetic-specific behavior flew in the face of the so-called unitary theory of general anesthesia, which maintains that all anesthetics work at the same site of action in all organisms (Deady et al., 1981; Table 1). unc-79 animals were first characterized as “fainters”, but otherwise moved well in air. Fainting refers to alternating periods of stopping and moving; motion is normal when moving but interrupted by periods of inactivity lasting several seconds (see Movie 2). unc-79 is not the only known mutation that confers a fainter phenotype. Animals carrying alleles of unc-80 (an uncharacterized gene; Morgan et al., 1988), and animals containing a loss of function mutation in both nca-1 (also known as unc-77 for gain-of-function alleles; E. Yeh, S. Ng and M. Zhen, personal communication) and nca-2, faint in air and are hypersensitive to anesthetics in a manner similar to unc-79 (K. Hamming, personal communication; Morgan and Sedensky, unpublished). By homology, both nca-1 and nca-2 encode probable cation channel proteins (Wormbase).
UNC-79 is a sentinel member of a novel family of highly conserved proteins expressed in the nervous systems of animals from nematodes to human. Expression of unc-79, which has 27 introns, appears to be tightly developmentally regulated when assayed by a GFP reporter. It is restricted to cells of the ventral nerve cord in young larvae, and becomes confined to a few head neurons in the adult. The gene has 3 introns which are highly conserved between C. elegans and C. briggsae, and its predominant message contains retained introns. It is likely that the expression pattern of unc-79 is dependent on which introns are retained. In unc-79 mutants, neither NCA-1 nor NCA-2 is found on Western blots, while transcription of these genes is normal (Sedensky and Morgan, unpublished). Thus, unc-79 may exert its effect through stabilization of these two proteins.
This behavior of unc-79 and unc-80 animals was completely suppressed by null mutations in the X-linked gene unc-1, which encodes the protein stomatin (Morgan et al., 1990; Rajaram et al., 1998). Animals containing different alleles of unc-1 display different behaviors in the gaseous anesthetics; unc-1(0) animals have a kinking motion and are only sensitive to diethyl ether, while unc-1(d) animals are hypersensitive to halothane, and unc-79;unc-1(d) animals are extremely anesthetic sensitive. The other kinked mutations unc-7, unc-9 and unc-24 also suppress unc-79, unc-80, and nca-2;nca-1 (Morgan and Sedensky, unpublished; Table 1).
unc-1 encodes a stomatin homologue that is broadly, but exclusively, expressed within the nematode nervous system (and in the 4 vulval muscles) in a pattern consistent with localization in the nerve cell membrane (Rajaram et al., 1998; Rajaram et al., 1999; Sedensky et al., 2001). Stomatins are membrane proteins that are associated with membrane channels, within microdomains known as lipid rafts (Snyers et al., 1999). Two other stomatins have been studied in C. elegans, UNC-24, which controls the distribution of UNC-1 (Sedensky et al., 2004) and MEC-2 which is involved with mechanosensation (Huang et al., 1995) and does not affect anesthetic sensitivity (Morgan and Sedensky, unpublished).
UNC-1 had been postulated to form homo-oligomers based on its complicated intra-allelic complementation patterns, a prediction now known to be true (Park and Horvitz, 1986; Snyers et al., 1998). In C. elegans it was established that UNC-1 was normally localized almost entirely within lipid rafts, and that it physically interacted with UNC-8, which is also primarily localized to rafts (Sedensky et al., 2004). unc-8 encodes a degenerin that is part of a large family of channel proteins that includes DEG/ENaCs (degenerins/epithelial sodium channels) and ASICs (acid-sensing ion channels; Tavernarakis et al., 1997; Waldmann et al., 1997). Like unc-1, unc-8 displays multiple classes of alleles (Park and Horvitz., 1986; Tavernarakis et al., 1997), which includes one with a phenotype similar to unc-1(0), i.e. a kinked phenotype that can suppress unc-79. unc-8(d);unc-1(d) displays a novel phenotype; this double mutant is a kinker unlike either coiler parent. UNC-1 and UNC-8 co-immunoprecipitate and thus probably physically interact in lipid rafts. Thus, there is a complicated interaction between these two gene products (Rajaram et al., 1999; Sedensky et al., 2004). Interestingly, UNC-79 is also apparently necessary for normal processing of UNC-8 within a raft, although UNC-79 itself appears to be confined to the cytosol (Morgan and Sedensky, unpublished).
It is interesting that the members of the SNARE complex have been found to be associated with lipid rafts and with ENaC channels (Lafont et al., 1999). Syntaxin mutations do not alter anesthetic sensitivity when immobility is used as an endpoint. However, the association of the SNARE complex, stomatin and ENaC channels with lipid rafts and with each other suggests a common pathway or target may be affected by these mutations. These data are further emphasized by the fact that the anesthetic response of yeast is modulated by the ubiquitin system, which also resides in lipid rafts (Wolfe et al. 1999; Lafont and Simons, 2001). Thus, proteins identified by diverse screens for changes in volatile anesthetic sensitivity may implicate related targets associated with lipid rafts in both yeast and in C. elegans.
Stomatins and ENaCs are known to co-localize within tissues like the dorsal root ganglion, part of the sensory pathway in mammals (Fricke et al., 2000). In addition, stomatins are known to affect channel properties of ASICs/ENaCs (Price et al., 2004). A mouse knock out for stomatin gives a profile of anesthetic sensitivity similar to that seen in the worm (Sedensky et al., 2006). This knockout is of a stomatin that is expressed in the spinal cord and periphery of the animal; there are no other mutant forms of stomatin for mammals (Zhu et al., 1999). In particular there is none specific to brain stomatin, and no dominant mutations similar to unc-1(d).
The above studies thus identify several genes that interact to affect sensitivity to volatile anesthetics. The identified genes include three that code for ion channels (unc-8, nca-1, and nca-2), two that encode stomatin-like proteins (unc-1, unc-24), two that encode gap junction proteins (unc-7, unc-9) and one novel protein (unc-79). The protein product of unc-80 remains to be identified. The interactions of these genes, as well as those that effect endpoints other than immobility, are summarized in Table 1.
Another mutation was identified in a screen for hypersensitivity to the volatile anesthetic isoflurane. Unlike unc-79 animals, this new mutation conferred extreme hypersensitivity to all volatile anesthetics, as well as to ethanol (Morgan and Sedensky, 1994; Kayser et al., 2003). Named gas-1 (general anesthetic sensitive), it is a temperature-sensitive lethal with reduced fecundity, increased oxygen sensitivity, and a shortened lifespan at 20°C. gas-1 encodes a subunit - an ortholog of the bovine “49kD” protein of complex I-, of the respiratory chain in mitochondria (Kayser et al., 1999). In the mitochondrion, electrons are passed from specific electron donors through a series of protein complexes to the final electron acceptor, oxygen (see Figure 2). A proton gradient is established in the process, which drives the synthesis of ATP. The concerted action of electron transport and ATP production is termed oxidative phosphorylation. Complex I is one of the points of entry for electrons into the respiratory chain and one of the proton pumps. It is the largest and least well-characterized complex, with an estimated size in mammals of almost 1000kD. (Hirst et al., 2003) gas-1 mitochondria are quite defective in oxidative phosphorylation using complex I specific substrates; individual steps of electron passage within the complex are also reduced, but rates of passage of electrons to CoQ is reduced by over 90% (Kayser et al., 2001). clk-1, a defect in CoQ synthesis, is also specifically defective in passage of electrons from complex I, and is mildly hypersensitive to volatile anesthetics (Hartman et al., 2001; Kayser et al., 2004a). RNAi induced hypomorphs in complex I subunits are also anesthetic sensitive, in a manner that matches their rates of oxidative phosphorylation with complex I substrates (Kayser et al., 2004b; Falk et al., 2006).
|
Figure 2. The integration of steps necessary for oxidative phosphorylation. Substrates that are electron donors for the respiratory chain are transported across the inner mitochondrial membrane by different carriers. In the matrix, electrons from pyruvate, glutamate, and malate are transferred to NAD+ by substrate-specific dehydrogenases and enter the respiratory chain via complex I. Succinate dehydrogenase is an integral activity of complex II. Electrons from complex I and II are transferred to complex III by the common shuttle CoQ. The electrons reach oxygen, the terminal acceptor, via cytochrome c and complex IV. Electron transport through the respiratory chain (flat gray arrows) is absolutely linked to proton extrusion by complexes I,-III, and IV (flat white arrows). The proton gradient across the inner mitochondrial membrane drives ATP production. ATP synthetase allows protons to re-enter the matrix and uses the energy released in this process to synthesize ATP. The substrates for phosphorylation, ADP and inorganic phosphate (Pi), enter the matrix via the ADP/ATP translocase (ANT) and the phosphate carrier. Measuring respiration of isolated mitochondria in response to natural and artificial electron donor substrates allows the integrated function of mitochondria to be assessed and the localization of defective steps. The positions of the defective steps for gas-1 (complex I), mev-1(complex II), isp-1(complex III), and clk-1(CoQ) mitochondria are indicated by the red bars.
In addition, daf-2 animals were tested for anesthetic sensitivity based on the longevity phenotype they share with clk-1. daf-2 animals are resistant to volatile anesthetics and have an increased rate of complex I dependent oxidative phosphorylation (Morgan and Sedensky, unpublished). The effects of daf-16 on this response have not been determined. In contrast, animals containing mutations in complex II (mev-1) and complex III (isp-1), which possess defects in other components of the electron transport chain, behave like wildtype in anesthetics (Kayser et al., 2004b; Falk et al., 2006). isp-1 animals have decreased rates of complex I function, as well as decreased rates of complex II, due to its position downstream in the electron transport chain (see Figure 2). Despite the decreased complex I dependent oxidative phosphorylation rates seen in isp-1, the anesthetic sensitivity of this mutant is normal. Previously, work had shown that radiolabeled halothane accumulates in mitochondria, with more label within the organelle in unc-79 animals than in N2. (R.E. Eckenhoff, personal communication) In mammals a similar labeling experiment showed that in the 25 proteins that could be identified as labeled with halothane, 17 were mitochondrial proteins. (Xi et al., 2004) It has also been demonstrated in mammals and in worms that of all the complexes of the electron transport chain, complex I is the most sensitive to inhibition by the volatile anesthetics (Kayser et al., 2004b; Cohen, 1973).
In the nematode, it does not appear to be amounts of ATP that account for the difference in anesthetic sensitivity between wildtype and mutant worms. Nor does oxidative damage of a putative anesthetic target explain the sensitivity of complex I mutants to volatile agents. The sum of data indicates that complex I rates of oxidative phosphorylation, assayed in vitro for near maximal capacity, reflect vulnerability of the animal to volatile anesthetics when the defect is due to a change within complex I itself. Defects in other complexes, or defects downstream of complex I, cause minor or no changes in anesthetic response. Mitochondrial function represents a novel potential mechanism of action of volatile anesthetics.
Regardless of the exact mechanism by which this novel putative target affects anesthetic behavior, a striking correlate of this finding has been found in patients (Morgan et al., 2002). Children undergoing surgery, primarily for muscle biopsies taken for diagnosis of suspected mitochondrial defects, were anesthetized with the volatile anesthetic sevoflurane (Morgan et al., 2002). Each patient's anesthetic sensitivity was determined via an integrated encephalogram obtained with a cutaneously applied sensor called a bispectral index (BIS). Those children with a confirmed complex I defect lost consciousness (indicated by a BIS reading of 60) at lower concentrations of anesthetic than normal children or those with other mitochondrial disorders - often by an order of magnitude.
At first glance data from C. elegans implicate quite disparate molecules as possible targets for volatile anesthetics. However, closer inspection indicates that these studies are beginning to form a picture of the molecular sites that control anesthetic sensitivity (see Figure 3). The genetic studies strongly suggest that anesthetic response is dependent on a broad group of molecular targets or pathways. The finding of altered sensitivity in patients with mitochondrial disease, the similarities between the responses of related mutants in mouse and nematodes, and the potential involvement of presynaptic proteins from mammals in anesthetic action underscores the applicability of these studies to the human population. A recent resurgence in interest in the role of mitochondria in regulating synaptic function may eventually tie a novel potential target to pathways that intuitively appear more directly related to the anesthetic state.
|
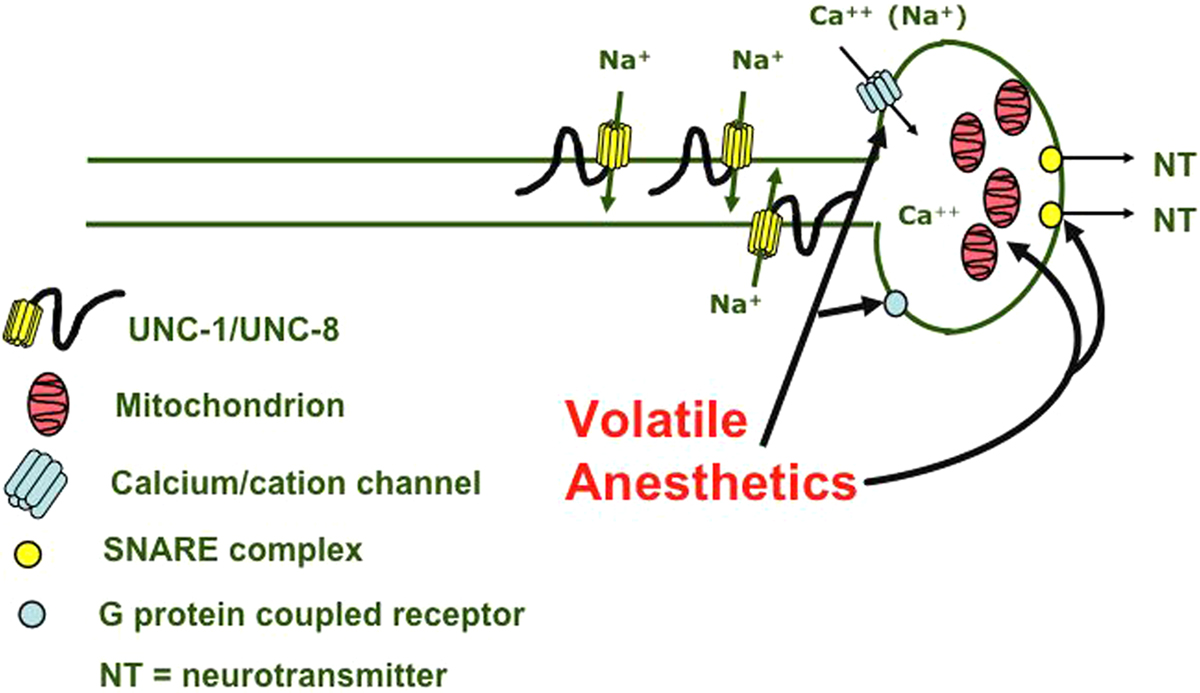
Figure 3. Model for volatile anesthetic function in the presynapse in the nematode. Arrows point to putative targets, which include the SNARE complex, mitochondria, G-protein coupled receptors, and cation channels. In addition, the genes unc-1 and unc-8 affect sensitivity, implicating the degenerin/ENaC family in the anesthetic response. These genes generally implicate the presynapse more strongly than the postsynapse as a major site of anesthetic action.
It is also interesting to note what genetic data fail to do. They do not identify a single protein or channel that is uniquely responsible for the effects of anesthetics. The many endpoints associated with the anesthetic state are probably the result of effects at different sites of anesthetic action. In addition, even when considering a defined simple endpoint, the volatile anesthetics behave differently from each other. This indicates that, even with single endpoints, multiple targets or mechanisms are involved. However, the recurring theme of presynaptic function affecting volatile anesthetic sensitivity makes this cellular region an enticing possibility.
It is clear that volatile anesthetics are invaluable, time-tested, tools of modern clinical medicine. Understanding the mechanism of volatile anesthetic action will certainly be the key to produce the perfect anesthetic. But many questions remain as to the basis of their clinical effect, some of which remain with us from the inception of their use. Are there truly multiple targets for volatile anesthetics, or are we measuring multiple effects upstream or downstream from a single anesthetic target? If there are multiple targets, then how many? Do they encompass a family of receptors, or very different sorts of molecules? How can the target(s) react with so many different types of molecules?
In addition, as a class of drugs that quickly and reversibly produce loss of consciousness and pain, volatile anesthetics represent potential experimental probes into large questions of modern biology. What is the mechanism of consciousness? The nature of normal sleep? Can novel means be found to treat acute and chronic pain? What is the relation between hibernation and the anesthetized state? If one assumes that fundamental physiologic processes are highly conserved within the animal kingdom, then using C. elegans as a model to understand the mechanism of action of volatile anesthetics is a logical starting point to answer these questions. Concurrently, significant efforts can be made to extrapolate findings in worms to more complex animals. Ultimate success in understanding how the volatile anesthetics work will have far reaching ramifications for modern biology.
Abraham, M.H., Lieb, W.R., and Franks, N.P. (1991). Role of hydrogen bonding in general anesthesia. J. Pharm. Sci. 80, 1309–1314. Abstract Article
Cohen, P.J. (1973). Effect of anesthetics on mitochondrial function. Anesthesiology 39, 153–164. Abstract Article
Crowder, C.M., Shebester, L.D., Schedl, T. (1996). Behavioral effects of volatile anesthetics in Caenorhabditis elegans. Anesthesiology 85, 901–912. Abstract Article
Davies, A.G., Pierce-Shimomura, J.T., Kim, H., VanHoven, M.K., Thiele, T.R., Bonci, A., Bargmann, C.I., and McIntire, S.L. (2003). A central role of the BK potassium channel in behavioral responses to ethanol in C. elegans. Cell 115, 655–666. Abstract Article
Deady, J.E., Koblin, D.D., Eger, E.I., II, Heavner, J.E., and D'Aoust, B. (1981). Anesthetic potencies and the unitary theory of narcosis. Anesth. Analg. 60, 380–384. Abstract Article
Evers, A.S., and Crowder, C.M. (2006). Cellular and molecular mechanisms of anesthesia. In: Clinical Anesthesia, P.G. Barash, B.F. Cullen, R.K. Stoelting, eds. (Lippincott Williams and Wilkins), pp 111–132.
Falk, M.J., Kayser, E.B., Morgan, P.G., and Sedensky, M.M. (2006). Mitochondrial complex I function modulates volatile anesthetic sensitivity in C. elegans. Curr. Biol. 16, 1641–1615. Abstract Article
Fricke, B., Lints, R., Stewart, G., et al. (2000). Epithelial Na+ channels and stomatin are expressed in rat trigeminal mechanosensory neurons. Cell Tissue Res. 299, 327–334. Abstract
Hartman, P.S., Ishii, N., Kayser, E.B., Morgan, P.G., and Sedensky, M.M. (2001). Mitochondrial Mutations Differentially Affect Aging, Mutability and Anesthetic Sensitivity in C. elegans. Mech. Ageing Dev. 122, 1187–1201. Abstract Article
Hawasli, A.H., Saifee, O., Liu, C., Nonet, M.L., and Crowder, C.M. (2004). Resistance to volatile anesthetics by mutations enhancing excitatory neurotransmitter release in Caenorhabditis elegans. Genetics 168, 831–843. Abstract Article
Hirst, J., Carroll, J., Fearnley, I.M., Shannon, R.J., and Walker, J.E. (2003). The nuclear encoded subunits of complex I from bovine heart mitochondria. Biochim. Biophys. Acta 1604, 135–150. Abstract Article
Huang, M., Gu, G., Ferguson, E.L., and Chalfie, M. (1995). A stomatin-like protein necessary for mechanosensation in C. elegans. Nature 378, 292–295. Abstract Article
Kayser, E.B., Hoppel, C.L., Morgan, P.G., and Sedensky, M.M. (2003). A mutation in mitochondrial complex I increases ethanol sensitivity in Caenorhabditis elegans. Alcohol. Clin. Exp. Res. 27, 584–592. Abstract Article
Kayser, E.B., Morgan, P.G., Hoppel, C.L., and Sedensky, M.M. (2001). Mitochondrial Expression and Function of GAS-1 in Caenorhabditis elegans. J. Biol. Chem. 276, 20551–20558. Abstract Article
Kayser, E.B., Morgan, P.G., Sedensky, M.M., and Hoppel, C.L. (2004a). The long-lived mutant clk-1 has a novel defect in mitochondrial oxidative phosphorylation. J. Biol. Chem. 279, 54479–54486. Abstract Article
Kayser, E.B., Morgan, P.G., and Sedensky, M.M. (2004b). Mitochondrial complex I function affects halothane sensitivity in Caenorhabditis elegans. Anesthesiology 101, 365–372. Abstract Article
Kayser, E.B., Morgan, P.G., and Sedensky, M.M. (1999). GAS-1, a mitochondrial protein controls sensitivity to volatile anesthetics in the nematode, C. elegans. Anesthesiology 90, 545–554. Abstract Article
Kayser, E.B., Rajaram, S., Thomas, S., Morgan, P.G., and Sedensky, M.M. (1998). Control of anesthetic response in C. elegans. Toxicol Lett. 100, 339–346. Abstract Article
Koushika, S.P. and Nonet, M.L. (2000). Sorting and transport in C. elegans: A model system with a sequenced genome. Curr. Opin. Cell Biol. 12, 517–523. Abstract Article
Lackner, M.R., Nurrish, S.J., and Kaplan, J.M. (1999). Facilitation of synaptic transmission by EGL-30 Gqalpha and EGL-8 PLCbeta: DAG binding to UNC-13 is required to stimulate acetylcholine release. Neuron 24, 335–346. Abstract Article
Lafont, F., and Simons, K. (2001). Raft-partitioning of the ubiquitin ligases Cbl and Nedd4 upon IgE-triggered cell signaling. Proc. Natl. Acad. Sci. U.S.A. 98, 3180–3184. Abstract Article
Lafont, F., Verkade, P., Galli, T., Wimner, C., Louvard, D., and Simons, K. (1999). Raft association of SNAP receptors acting in apical trafficking in Madin-Darby canine kidney cells. Proc. Natl. Acad. Sci. U.S.A. 96, 3734–3738. Abstract Article
Meyer, H.H. (1899). Zur Theorie der alkoholnarkose. I. Mitt. Welche eigenschaft der anasthetika bedingt ihre narkotische wirkung. Arch. Exp. Pathol. Pharmak. 42, 109–119. Article
Miller, K.G., Emerson, M.D., and Rand, J.B. (1999). Goalpha and diacylglycerol kinase negatively regulate the Gqalpha pathway in C. elegans. Neuron 24, 323–333. Abstract Article
Morgan, P.G., Hoppel, C.L., and Sedensky, M.M. (2002). Mitochondrial defects and anesthetic sensitivity. Anesthesiology 96, 1268–1270. Abstract Article
Morgan, P.G., Sedensky, M.M., Meneely, P.M., and Cascorbi, H.F. (1988). The effect of two genes on anesthetic response in the nematode Caenorhabditis elegans. Anesthesiology 69, 246–251. Abstract Article
Morgan, P.G., Sedensky, M.M., and Meneely, P.M. (1990). Multiple sites of action of volatile anesthetics in Caenorhabditis elegans. Proc. Natl. Acad. Sci. U.S.A. 87, 2965–2969. Abstract Article
Morgan, P.G., and Sedensky, M.M. (1994). Mutations conferring new patterns of sensitivity to volatile anesthetics in Caenorhabditis elegans. Anesthesiology 81, 888–898. Abstract Article
Morgan, P.G. and Cascorbi, H.F. (1985). Effect of anesthetics and a convulsant on normal and mutant Caenorhabditis elegans. Anesthesiology 62, 738–744. Abstract Article
Moss, G.W., Franks, N.P., and Lieb, W.R. (1991). Modulation of the general anesthetic sensitivity of a protein: a transition between two forms of firefly luciferase. Proc. Natl. Acad. Sci. U.S.A. 88, 134–138. Abstract Article
Nagele, P., Mendel, J.B., Placzek, W.H., Scott, B.A., d'Avignon, D.A., and Crowder, C.M. (2005a). Volatile anesthetics bind rat synaptic snare proteins. Anesthesiology 103, 768–778. Abstract Article
Nagele, P., Metz, L.B., and Crowder, C.M. (2005b). Xenon acts by inhibition of non-N-methyl-D-aspartate receptor-mediated glutamatergic neurotransmission in Caenorhabditis elegans. Anesthesiology 103, 508–513. Abstract Article
Nagele, P., Metz, L.B., and Crowder, C.M. (2004). Nitrous oxide requires the N-methl-D-aspartate receptor for its action in Caenorhabditis elegans. Proc. Natl. Acad. Sci. U.S.A. 101, 8791–8796. Abstract Article
Nonet, M.L., Saifee, O., Zhao, H., Rand, J.B., and Wei, L. (1998). Synaptic transmission deficits in Caenorhabditis elegans synaptobrevin mutants. J. Neurosci. 18, 70–80. Abstract
Park, E.C., and Horvitz, H.R. (1986). Mutations with dominant effects on the behavior and morphology of the nematode Caenorhabditis elegans. Genetics 113, 821–852. Abstract
Price, M.P., Thompson, R.J., Eshcol, J.O., Wemmie, J.A., and Benson, C.J. (2004). Stomatin modulates gating of acid-sensing ion channels. J. Biol. Chem. 279, 53886–53891. Abstract Article
Rajaram, S., Sedensky, M.M., and Morgan, P.G. (1998). unc-1, a stomatin homologue controls sensitivity to volatile anesthetics in C. elegans. Proc. Natl. Acad. Sci. U.S.A. 95, 8761–8766. Abstract Article
Rajaram, S., Spangler, T.L., Sedensky, M.M., and Morgan, P.G. (1999). A stomatin and a degenerin interact to control anesthetic sensitivity in C. elegans. Genetics 153, 1673–1682. Abstract
Rand, J.B., and Nonet, M.L. (1997). Synaptic transmission. In: Riddle, D.L., Blumenthal, T., Meyer, B.J. and Priess, J.R. (eds), C. elegans II. (Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press), pp. 611–643.
Regan, M.J., Eger, E.I., 2nd. (1967). Effect of hypothermia in dogs on anesthetizing and apneic doses of inhalation agents. Determination of the anesthetic index (Apnea/MAC). Anesthesiology 28, 689–700. Abstract Article
Sedensky, M.M., and Meneely, P.M. (1987). Genetic analysis of halothane sensitivity in Caenorhabditis elegans. Science 236, 952–954. Abstract Article
Sedensky, M.M., Siefker, J., and Morgan, P.G. (2001). Model organisms: new insights into ion channel and transporter function. Stomatin homologues interact in C. elegans. Am. J. Physiol., Cell Physiol. 280, C1340–C1348. Abstract
Sedensky, M.M., Siefker, J.M., and Morgan, P.G. (2004). A stomatin and a degenerin interact in lipid rafts of the nervous system of Caenorhabditis elegans. Am. J. Physiol., Cell Physiol. 287, C468–C474. Abstract Article
Sedensky, M.M., Pujazon, M.A., and Morgan, P.G. (2006). Tail clamp responses in stomatin knockout mice compared with mobility assays in Caenorhabditis elegans during exposure to diethyl ether, halothane, and isoflurane. Anesthesiology 105, 498–502. Abstract Article
Snyers, L., Umlauf, E., and Prohaska, R. (1999). Association of stomatin with lipid-protein complexes in the plasma membrane and the endocytic compartment. Eur. J. Cell Biol. 78, 802–812. Abstract
Snyers, L., Umlauf, E., and Prohaska, R. (1998). Oligomeric nature of the integral membrane protein stomatin. J. Biol. Chem. 273, 17221–17226. Abstract Article
Suezaki, Y., Tatara, T., Kaminoh, Y., Kamaya, H., and Ueda, I. (1990). A solid-solution theory of anesthetic interaction with lipid membranes: temperature span of the main phase transition. Biochim. Biophys. Acta 1029, 143–148. Abstract Article
Tavernarakis, N., Shreffler, W., Wang, S., and Driscoll, M. (1997). unc-8, a DEG/ENaC family member, encodes a subunit of a candidate mechanically gated channel that modulates C. elegans locomotion. Neuron 18, 107–119. Abstract Article
Tanifuji, Y., Eger, E.I., II, and Terrell, R.C. (1977). Some characteristics of an exceptionally potent inhaled anesthetic: thiomethoxyflurane. Anesth. Analg. 56, 387–390. Abstract Article
van Swinderen, B., Metz, L.B., Shebester, L.D., and Crowder, C.M. (2002). A Caenorhabditis elegans pheromone antagonizes volatile anesthetic action through a go-coupled pathway. Genetics 161, 109–119. Abstract
van Swinderen, B., Metz, L.B., Shebester, L.D., Mendel, J.E., Sternberg, P.W., and Crowder, C.M. (2001). Goalpha regulates volatile anesthetic action in Caenorhabditis elegans. Genetics 158, 643–655. Abstract
van Swinderen, B., Saifee, O., Shebester, L., Roberson, R., Nonet, M.L., and Crowder, C.M. (1999). A neomorphic syntaxin mutation blocks volatile-anesthetic action in Caenorhabditis elegans. Proc. Natl Acad. Sci. U.S.A. 96, 2479–2484. Abstract Article
Waldmann, R., Champigny, G., Bassilana, F., Heurteaux, C., and Lazdunski, M. (1997). A proton-gated cation channel involved in acid-sensing. Nature 386, 173–177. Abstract Article
Wang, Z.W., Saifee, O., Nonet, M.L., and Salkoff, L. (2001). SLO-1 potassium channels control quantal content of neurotransmitter release at the C. elegans neuromuscular junction. Neuron 32, 867–881. Abstract Article
Wolfe, D., Reiner, T., Keeley, J.L., Pizzini, M., and Keil, R.L. (1999). Ubiquitin metabolism affects cellular response to volatile anesthetics in yeast. Mol. Cell. Biol. 19, 8254–8262. Abstract
Wu, X.-S., Sun, J.-Y., Evers, A.S., Crowder, M., and Wu, L.G. (2004). Isoflurane inhibits transmitter release and the presynaptic action potential. Anesthesiology 100, 663–700. Abstract Article
Xi, J., Liu, R., Asbury, G.R., Eckenhoff, M.F., and Eckenhoff, R.G. (2004). Inhalational anesthetic-binding proteins in rat neuronal membranes. J. Biol. Chem. 279, 19628–19633. Abstract Article
Zhang, Y., Trudell, J.R., Mascia, M.P., Laster, M.J., Gong, D.H., Harris, R.A., Eger, E.I., II. (2000). The anesthetic potencies of alkanethiols for rats: relevance to theories of narcosis. Anesth. Analg. 91, 1294–1299. Abstract Article
Zhu, Y., Paszty, C., Turetsky, T., Tsai, S., Kuypers, F.A., Lee, G., Cooper, P., Gallagher, P.G., Stevens, M.E., Rubin, E., Mohandas, N., and Mentzer, W.C. (1999). Stomatocytosis is absent in “stomatin”-deficient murine red blood cells. Blood 93, 2404–2410. Abstract
*Edited by Adres Villu Maricq and Steven McIntire. Last revised December 28, 2006. Published May 3, 2007. This chapter should be cited as: Morgan, P.G., et al. C. elegans and volatile anesthetics (May 3, 2007), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.140.1, http://www.wormbook.org.
Copyright: © 2007 P.G. Morgan, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
§To whom correspondence should be addressed. E-mail: margaret.sedensky@uhhs.com
 All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.
All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.