Embryo series courtesy of Einhard Schierenberg
Embryo series courtesy of Einhard SchierenbergTable of Contents
Abstract
In response to harsh environmental conditions, C. elegans larvae undergo dauer arrest at the second molt. The past decade has yielded many insights into the signaling pathways and the molecular mechanisms that govern this developmental transition. Dauer pheromone, the major physiologic signal promoting dauer arrest, has been purified, identified, and synthesized. The molecular identities of the vast majority of dauer regulatory genes isolated in initial genetic screens are now known. Physiologic ligands for DAF-12, a nuclear receptor that is the final common target of dauer regulatory pathways, have been identified. The discovery of the Hid (high temperature induction of dauer) phenotype and the results of enhancer screens have greatly expanded the repertoire of dauer regulatory genes. Genomic analysis of dauer arrest has highlighted the role of pathway crosstalk in dauer regulation. Nonetheless, critical questions remain about the mechanistic underpinnings of dauer arrest.
In 1975, Cassada and Russell described the dauerlarva, an arrested developmental variant of C. elegans that forms at the second molt in response to environmental duress. Dauer larvae exhibit morphological and behavioral characteristics distinct from those of larvae developing in replete environments (Cassada and Russell, 1975). Since that discovery, the transition to dauer arrest has been a fertile area of investigation in the C. elegans field, both as a model for regulatory mechanisms governing morphological change during organismal development as well as a parallel to obligate dauer-like developmental stages in parasitic nematodes. In recent years, the molecular identities of genes mutated in various mutants exhibiting dysregulation of dauer arrest (daf mutants) have revealed the critical roles of evolutionarily conserved guanylyl cyclase, transforming growth factor-β (TGFβ)-like, insulin-like, and hormonal signaling pathways in the regulation of dauer arrest.
The dauer developmental decision occurs late in the L1 larval stage and hinges on the integration of three environmental parameters: population density, food supply, and ambient temperature. Population density is the major influence on dauer arrest and is conveyed by a constitutively synthesized pheromone that induces dauer arrest and prevents dauer recovery. Conversely, a heat-stable, hydrophilic “food signal” that provides information about food availability inhibits dauer arrest and promotes dauer recovery. The relative amounts of pheromone and food signal, rather than absolute levels, are critical in determining whether an animal undergoes dauer arrest (Golden and Riddle, 1982). Increased temperature enhances pheromone-induced dauer arrest (Golden and Riddle, 1984a), and some temperature-sensitive dauer-constitutive mutants are suppressible by an amber nonsense suppressor (Golden and Riddle, 1984b), suggesting that dauer arrest is intrinsically dependent upon ambient temperature.
Dauer larvae are morphologically distinct from larvae that develop in replete conditions. Dauers are radially constricted and possess a specialized cuticle with alae (Cassada and Russell, 1975). Their oral orifices are closed by an internal plug (Riddle et al., 1981), and their pharynxes are constricted (Vowels and Thomas, 1992) and do not pump (Cassada and Russell, 1975). Dauers are easily distinguishable from L3 larvae under a dissecting microscope. The dauer-specific cuticle and the lack of pharyngeal pumping confer resistance to many environmental insults, including 1% SDS (Cassada and Russell, 1975). SDS resistance provides a convenient method for isolating dauers and has greatly facilitated the genetic analysis of dauer formation.
C. elegans undergoes a metabolic shift between L1 and L2 stages; embryos and L1 larvae use the glyoxylate cycle to generate carbohydrates from lipid stores, whereas L2 and later stages shift toward aerobic respiration and exhibit a relative increase in TCA cycle activity. Dauers do not undergo this shift toward aerobic respiration (Wadsworth and Riddle, 1989). Relative to growing larvae, dauers also appear to be transcriptionally quiescent but have elevated levels of mRNA encoding the heat shock protein Hsp90 (Dalley and Golomb, 1992; Snutch and Baillie, 1983) and elevated superoxide dismutase (Larsen, 1993; Vanfleteren and De Vreese, 1995) and catalase (Vanfleteren and De Vreese, 1995) activities. This suggests that dauers are adapted to resist metabolic stress and is consistent with the observation that they are long-lived (Klass and Hirsh, 1976).
Dauer pheromone has recently been purified from C. elegans extracts and molecularly characterized. Paik and colleagues first demonstrated that the ascaroside (-)-6-(3,5-dihydroxy-6-methyltetrahydropyran-2-yloxy) heptanoic acid has dauer pheromone activity (Jeong et al., 2005). Recently, Jon Clardy's group has identified three distinct ascarosides in C. elegans extracts with dauer pheromone activity (including the molecule identified by Paik's group). The two ascarosides distinct from that initially isolated by Paik's group, 5-O-ascarylosyl-5R-hydroxy-2-hexanone and an ascaroside derivative of 8R-hydroxy-2E-nonenoic acid, are two orders of magnitude more potent than (-)-6-(3,5-dihydroxy-6-methyltetrahydropyran-2-yloxy) heptanoic acid at inducing dauer arrest (Butcher et al., 2007). These discrepancies may be a consequence of differences in the purification steps used by the two groups (Jeong et al., 2005; Butcher et al., 2007). Furthermore, during purification, one group followed activity using a dauer formation assay (Jeong et al., 2005), whereas the other followed activity using a dauer recovery assay (Butcher et al., 2007).
Little is known about how dauer pheromone is synthesized in vivo and released into the environment. Culture medium and organic extracts of a dauer-defective mutant, daf-22(m130), do not contain dauer pheromone activity, suggesting that daf-22(m130) is defective in dauer pheromone synthesis (Golden and Riddle, 1985). Furthermore, exogenous dauer pheromone rescues the daf-22(m130) dauer-defective phenotype (Jeong et al., 2005). Interestingly, aqueous extracts of daf-22(m130) animals have measurable dauer pheromone activity (Golden and Riddle, 1985), suggesting that these animals may have a defect in a late step in dauer pheromone synthesis that results in the accumulation of a hydrophilic precursor or precursors that possess dauer pheromone activity. The molecular identity of daf-22 is not known.
The mechanisms underlying direct cellular responses to dauer pheromone have not been elucidated. It has been assumed that dauer pheromone binds to a specific cell surface receptor expressed in amphid neurons; however, the molecular identity of such a receptor has remained elusive. The demonstration that active dauer pheromone consists of multiple compounds (Butcher et al., 2007) suggests that each compound may bind to a distinct receptor that transduces a dauer-promoting signal. This may explain why candidate dauer pheromone receptors have not emerged from the extensive genetic analysis of dauer development that has been performed to date. The availability of synthetic pheromone components should facilitate the search for their cognate receptors.
Consistent with the possibility that dauer pheromone binds to and activates a G-protein-coupled seven-transmembrane domain receptor, the G-protein α-subunits GPA-2 and GPA-3 promote dauer arrest when activated, suppress pheromone-induced dauer arrest when deleted, and are expressed in amphid neurons (Zwaal et al., 1997). The ability of cyclic GMP (cGMP) analogs to suppress pheromone-induced dauer arrest (Birnby et al., 2000) suggests that dauer pheromone may cause dauer arrest at least in part by antagonizing a guanylyl cyclase. Many transmembrane guanylyl cyclase receptors are expressed in amphid neurons (Birnby et al., 2000; Yu et al., 1997) and could potentially regulate responses to dauer pheromone.
Genetic analysis of dauer-constitutive (Daf-c) and dauer-defective (Daf-d) mutants has defined four distinct pathways that regulate dauer arrest (Gerisch et al., 2001; Gottlieb and Ruvkun, 1994; Jia et al., 2002; Riddle et al., 1981; Thomas et al., 1993; Vowels and Thomas, 1992). In the past decade, components of these and other pathways have been cloned and characterized, revealing the complex interplay of at least four evolutionarily conserved signal transduction pathways in regulating dauer formation.
The Daf-c gene daf-11 defines a pathway that inhibits dauer arrest in parallel (at least in part) to a pathway defined by the genes daf-1, 4, 7, 8, and 14 (Thomas et al., 1993). daf-11 mutants also have defects in chemosensation (Vowels and Thomas, 1994). daf-11 encodes a transmembrane guanylyl cyclase that is expressed in the chemosensory neurons ASI, ASJ, ASK, AWB, and AWC (Birnby et al., 2000). 8-bromo-cGMP rescues the Daf-c phenotype of daf-11 mutants, suggesting that the Daf-c phenotype of daf-11 mutants is secondary to a reduction in cGMP synthesis. The inability of 8-bromo-cGMP to rescue dauer arrest in most other Daf-c mutants suggests that DAF-11 functions proximally in the regulation of dauer arrest (Birnby et al., 2000).
Components of a cGMP-gated ion channel are candidate downstream effectors of DAF-11. tax-2 and tax-4 encode subunits of a cGMP-gated ion channel that regulates thermosensation, chemosensation, and neuronal development (Coburn and Bargmann, 1996; Coburn et al., 1998; Komatsu et al., 1996). Both tax-2 and tax-4 are expressed in sensory neurons and have overlapping expression patterns with daf-11, indicating that they may function together in the same cells. The tax-4 dauer arrest phenotype is not rescued by 8-bromo-cGMP (Birnby et al., 2000), suggesting that TAX-4 is a target of DAF-11-dependent cGMP signaling. These data suggest that DAF-11 inhibits dauer arrest at least in part by activating TAX-2 and TAX-4 through increased cGMP synthesis. tax-4 mutants have a weaker Daf-c phenotype than daf-11 mutants (Coburn et al., 1998), indicating that DAF-11 probably activates multiple downstream effectors that contribute to inhibition of dauer arrest.
The daf-21 gene was defined by a single allele, p673, that has genetic interactions and chemosensory pleiotropies similar to those of daf-11 mutants (Thomas et al., 1993; Vowels and Thomas, 1994). Detailed genetic analysis of daf-21(p673) indicates that it is a recessive gain-of-function allele and that its phenotypes may be a consequence of allele-specific inhibition of DAF-11 function. Consistent with this finding, a null allele of daf-21 does not have a dauer arrest phenotype and is larval lethal. daf-21 encodes a Hsp90 homolog (Birnby et al., 2000).
The TGFβ-like pathway that regulates dauer arrest is defined by the Daf-c genes daf-1, 4, 7, 8, and 14 and the Daf-d genes daf-3 and daf-5 (Patterson and Padgett, 2000). TGFβ family ligands bind to and activate heteromeric cell surface receptors with serine/threonine kinase activity, resulting in the phosphorylation and activation of SMAD transcription factors that translocate to the nucleus to regulate transcription (Shi and Massague, 2003). Previous reviews (Riddle and Albert, 1997) described the molecular characterization of homologs of TGFβ (DAF-7; Ren et al., 1996; Schackwitz et al., 1996) and Type I (DAF-1; Georgi et al., 1990) and Type II (DAF-4; Estevez et al., 1993) TGFβ receptors. Since then, cloning of the remaining genes in this pathway has defined downstream transcription factors that are targets of DAF-7/TGFβ-like signaling. This and other C. elegans TGFβ-like signal transduction pathways are described in detail in another section of WormBook (see TGFβ signaling). With the exception of DAF-7, which is expressed primarily in the ASI amphid neuron (Ren et al., 1996; Schackwitz et al., 1996), all components of this pathway are widely expressed (Da Graca et al., 2004; Gunther et al., 2000; Inoue and Thomas, 2000b; Patterson et al., 1997; Tewari et al., 2004). Mosaic analysis and rescue experiments with tissue-specific translational fusions indicate that DAF-4 functions non-autonomously to regulate dauer arrest (Inoue and Thomas, 2000b).
daf-3, daf-8, and daf-14 encode SMAD transcription factors (Estevez, 1997; Inoue and Thomas, 2000b; Patterson et al., 1997), and daf-5 encodes a Sno/Ski oncoprotein homolog that binds to DAF-3 (Da Graca et al., 2004; Tewari et al., 2004). Consistent with the role of mammalian SMADs as positive effectors of TGFβ-like signaling (Shi and Massague, 2003), DAF-8 and DAF-14 are required for DAF-7/TGFβ-mediated inhibition of dauer arrest (Inoue and Thomas, 2000b). However, in contrast, DAF-3 appears to be inhibited upon activation of the DAF-7/TGFβ pathway (Patterson et al., 1997).
Recent findings indicate that the role of DAF-3 in the regulation of dauer arrest may be more complex than initially appreciated. Although daf-3 loss-of-function mutations are Daf-d at 25°C (Thomas et al., 1993), they are Daf-c at 27°C (Ailion and Thomas, 2000), indicating that temperature may influence DAF-3 activity. Furthermore, although daf-3 mutants suppress 25°C dauer arrest in the Daf-c mutants daf-1, 4, 7, 8, and 14 (Thomas et al., 1993), they enhance the weak 25°C dauer arrest phenotype of some Daf-c mutants (Gerisch et al., 2001; Ohkura et al., 2003). Thus, DAF-3 can function to promote or to inhibit dauer arrest depending upon environmental and genetic context. The mechanisms underlying these observations remain obscure.
A genetic screen to identify suppressors of the dauer arrest phenotype of daf-1, daf-8, and daf-14 mutants identified three novel genes, scd-1, scd-2, and scd-3 (Scd: suppressors of constitutive dauer arrest) that have weak Daf-d phenotypes when mutated (Inoue and Thomas, 2000a). scd-1 and scd-2 appear to be novel components of the TGFβ pathway, whereas scd-3 suppresses both DAF-7/TGFβ pathway mutants as well as daf-11 mutants and may function downstream of both pathways (Inoue and Thomas, 2000a). scd-1 encodes a glutamine-rich protein, and scd-2 encodes a homolog of the mammalian anaplastic lymphoma kinase (WormBase website, http://www.wormbase.org, release WS170, February 10, 2007). SCD-2 inhibits synaptic differentiation and is antagonized by the F-box-containing synaptic protein FSN-1 (Liao et al., 2004). The molecular identity of scd-3 is not known.
egl-4 encodes a cGMP-dependent protein kinase that regulates chemosensation, locomotion, and body size (Daniels et al., 2000; Fujiwara et al., 2002; L'Etoile et al., 2002; Stansberry et al., 2001). egl-4 mutants are hypersensitive to dauer pheromone (Golden and Riddle, 1984b) and have a weak dauer arrest phenotype (Daniels et al., 2000). Although the molecular identity of egl-4 suggests that it may function in the guanylyl cyclase pathway to inhibit dauer arrest, genetic analysis indicates that egl-4 functions primarily in the TGFβ pathway to regulate dauer formation (Daniels et al., 2000).
BRA-1, a protein that binds to DAF-1/TGFβRI, appears to inhibit TGFβ signaling proximally, as a bra-1 deletion allele partially suppresses the dauer arrest phenotype of daf-1 mutants but does not suppress daf-14/SMAD dauer arrest (Morita et al., 2001). kin-8, which encodes a homolog of the ROR family of tyrosine kinases, promotes dauer bypass and is required for full expression of DAF-7/TGFβ in the ASI amphid neuron (Koga et al., 1999).
A third pathway regulating dauer formation was originally defined by the Daf-c genes daf-2 and daf-23 (allelic to age-1) and the Daf-d gene daf-16 and functions in parallel to the guanylyl cyclase and TGFβ-like pathways (Gottlieb and Ruvkun, 1994; Riddle and Albert, 1997; Thomas et al., 1993). Mutations affecting this pathway are phenotypically distinct from other Daf-c mutations in that strong alleles of daf-2 and age-1 arrest as dauers non-conditionally and do not recover (Gems et al., 1998; Morris et al., 1996). Furthermore, unlike guanylyl cyclase pathway and TGFβ pathway mutants, daf-2 pathway mutants exhibit extended lifespans (Friedman and Johnson, 1988; Hertweck et al., 2004; Kenyon et al., 1993; Morris et al., 1996; Paradis et al., 1999; Wolkow et al., 2002). The impact of this pathway on C. elegans longevity is described in detail in another WormBook chapter and will not be discussed further here.
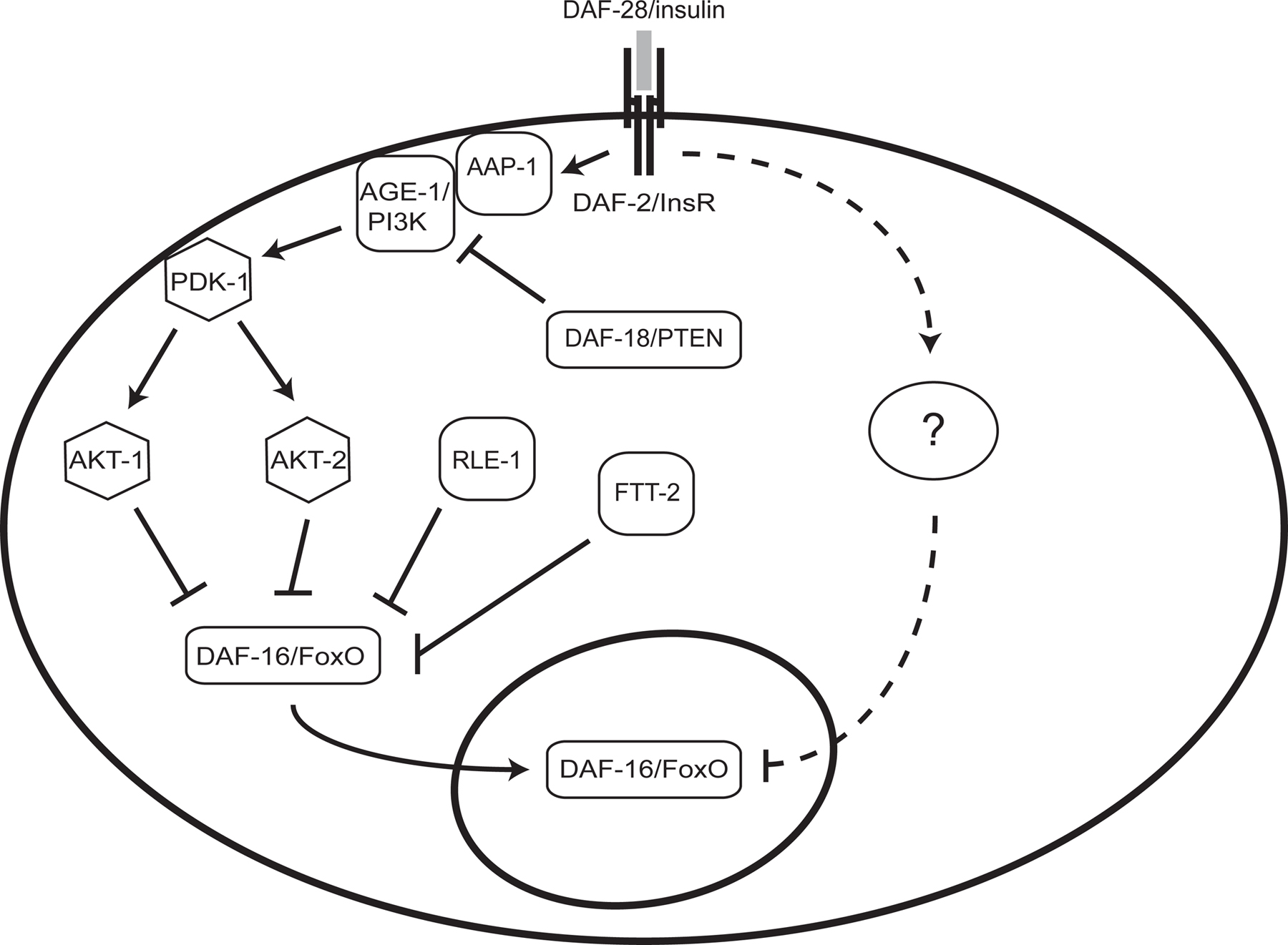
Cloning and characterization of these and other genes in this pathway have defined a conserved insulin-like pathway that inhibits dauer arrest through activation of the DAF-2 insulin receptor homolog, AGE-1 phosphoinositide 3-kinase (PI3K), and the protein kinases PDK-1, AKT-1, and AKT-2 (Kimura et al., 1997; Morris et al., 1996; Paradis et al., 1999; Paradis and Ruvkun, 1998). AKT-1 phosphorylates and inhibits the FoxO transcription factor DAF-16 by promoting its sequestration in the cytoplasm (Hertweck et al., 2004; Lee et al., 2001; Lin et al., 1997; Lin et al., 2001; Ogg et al., 1997). Cytoplasmic retention of DAF-16 requires the 14-3-3 proteins PAR-5 and FTT-2 (Berdichevsky et al., 2006; Li et al., 2007). DAF-18, the C. elegans ortholog of the phosphoinositide 3-phosphatase PTEN, antagonizes AGE-1/PI3K (Gil et al., 1999; Mihaylova et al., 1999; Ogg and Ruvkun, 1998; Rouault et al., 1999). The E3 ubiquitin ligase RLE-1 catalyzes polyubiquitination and proteasomal degradation of DAF-16 (Li et al., 2007; see Figure 1). As is the case for TGFβ pathway components, most of these genes are expressed throughout the animal. Mosaic analysis (Apfeld and Kenyon, 1998) and rescue experiments with tissue-specific translational fusions (Wolkow et al., 2000) indicate that DAF-2/InsR and AGE-1/PI3K function non-autonomously to inhibit dauer arrest.
Although most mutants with reduced insulin-like signaling have both Daf-c and extended longevity phenotypes, recent studies illustrate that dauer arrest and lifespan regulation by insulin-like signaling are decoupled by spatiotemporal compartmentalization. Intriguingly, although studies on DAF-2/InsR suggest that it functions primarily in the nervous system to regulate lifespan as well as dauer arrest (Apfeld and Kenyon, 1998; Wolkow et al., 2000), similar studies on the major target of insulin-like signaling, DAF-16/FoxO, suggest that dauer arrest and lifespan may be regulated by DAF-16/FoxO activity in distinct tissues. DAF-16/FoxO in the nervous system has stronger influences on dauer arrest than on lifespan, whereas intestinal DAF-16/FoxO plays a greater role in regulating lifespan than in regulating dauer arrest (Libina et al., 2003). Thus, DAF-16/FoxO activation in different tissues may have distinct phenotypic consequences. Furthermore, insulin-like signaling during larval development regulates dauer arrest without significantly impacting lifespan, whereas insulin-like signaling during adulthood suffices to regulate longevity (Dillin et al., 2002), indicating that temporal specificity also determines (at least in part) the biological output of insulin-like signaling.
Strikingly, the C. elegans genome is predicted to encode 40 insulin-like (INS) molecules (Pierce et al., 2001; WormBase website, http://www.wormbase.org, release WS170, February 10, 2007). Studies on ins genes are complicated by the potential functional redundancy of these genes as well as the possible indirect effects of INS protein overexpression on the processing of other INS family members. Notwithstanding these caveats, there is substantial evidence to indicate that ins genes regulate dauer arrest.
|
Figure 1. An insulin-like signaling pathway that regulates dauer arrest in C. elegans. See text for details.
ins-1 encodes an INS protein that is most similar to human insulin among C. elegans INS proteins. An ins-1 deletion mutant does not have a dauer arrest phenotype. However, overexpression of ins-1 or a cDNA encoding human insulin enhances dauer arrest, both in wild-type and daf-2/InsR mutant backgrounds. This suggests that INS-1 and human insulin antagonize DAF-2/InsR signaling. Although it remains unclear whether this effect is direct or indirect, there is some specificity among INS family members, as overexpression of ins-18 also enhanced dauer arrest, whereas overexpression of ins-9, ins-22, ins-19, or ins-31 did not. It is noteworthy that INS-1 and INS-18 are the only C. elegans INS proteins predicted to have C-peptides, which are proteolytically cleaved from proinsulin during normal insulin processing. Promoter::GFP fusions of many of these genes are expressed in amphid neurons, suggesting that dauer-regulatory environmental inputs could regulate expression and/or secretion of insulins (Pierce et al., 2001).
The Daf-c gene daf-28 encodes an INS family member that promotes insulin-like signaling. It is the only ins mutant that has emerged from forward genetic screens and is defined by a single semidominant allele (Li et al., 2003; Malone and Thomas, 1994). Strikingly, a daf-28p::GFP transgene is expressed in the amphid neurons ASI and ASJ and is downregulated in response to pheromone and starvation. The related INS proteins INS-4 and INS-6 appear to function similarly to DAF-28 to promote insulin-like signaling (Li et al., 2003). INS-6 can bind directly to and activate the human insulin receptor in vitro (Hua et al., 2003). A daf-28 deletion mutant does not undergo dauer arrest at 25°C (Weiqing Li, personal communication).
INS-7 may also promote reproductive development, as ins-7 RNAi enhances dauer arrest in a daf-2(e1370) mutant background (Murphy et al., 2003). Recently it has been shown that the ATPase ASNA-1 potentiates DAF-2/InsR signaling by promoting the secretion of INS peptides such as DAF-28 (Kao et al., 2007). The biological role of most of the other insulin-like proteins remains to be elucidated.
Orthologs of components of the conserved target-of-rapamycin (TOR) pathway that is activated by insulin-like molecules (Oldham and Hafen, 2003) also promote reproductive development in C. elegans. TOR is a protein kinase activated by insulin that couples global mRNA translation to nutrient availability. It exists in at least two heteromeric complexes, TORC1 and TORC2, which have distinct subunit compositions and biological functions. TORC1, which is sensitive to inhibition by rapamycin and contains the TOR binding partner Raptor, is required for activation of the ribosomal S6 kinase, whereas TORC2, which is insensitive to rapamycin and contains the TOR binding protein Rictor, phosphorylates and activates Akt (Inoki and Guan, 2006). let-363 and daf-15 encode orthologs of TOR and Raptor, respectively (Jia et al., 2004; Long et al., 2002). In contrast to loss-of-function mutants of daf-2, age-1, and other upstream components of insulin-like signaling, which arrest as complete dauers, let-363/TOR and daf-15/Raptor mutants arrest as dauer-like larvae that lack cuticular and intestinal characteristics of full dauers (Jia et al., 2004). This larval arrest phenotype is epistatic to daf-16/FoxO loss-of-function mutations, suggesting that LET-363/TOR and DAF-15/Raptor act downstream of or parallel to DAF-16/FoxO. In replete environments, insulin-like ligands may activate LET-363/TOR and DAF-15/Raptor to promote reproductive development. Inactivation of LET-363/TOR and DAF-15/Raptor suffices to induce developmental arrest, but other effectors of insulin-like signaling must also be inactivated in order for full dauer morphogenesis to take place.
Genetic evidence supports the existence of additional DAF-2 outputs that function in parallel to AGE-1/PI3K (see Figure 1). Activating mutations in akt-1 and pdk-1 suppress dauer arrest in age-1 null mutants but do not efficiently suppress dauer formation in daf-2(e1370) mutants (Inoue and Thomas, 2000a; Paradis et al., 1999; Paradis and Ruvkun, 1998), and the weak loss-of-function allele daf-18(e1375) suppresses dauer arrest in age-1 null mutants but not in daf-2(e1370) mutants (Gil et al., 1999; Gottlieb and Ruvkun, 1994; Inoue and Thomas, 2000a; Ogg and Ruvkun, 1998; Vowels and Thomas, 1992). Also, DAF-16/FoxO localized to the nucleus by virtue of either AKT-1 inactivation, inactivation of the 14-3-3 protein FTT-2, or mutation of consensus AKT-1 phosphorylation sites is not fully active unless daf-2 is also mutated (Berdichevsky et al., 2006; Hertweck et al., 2004; Lin et al., 2001). These results suggest that DAF-2 also inhibits the activity of nuclear DAF-16/FoxO in an AKT-independent manner. Candidate components of this parallel pathway have been isolated recently (described below in the section on modifier screens).
Although genetic analysis is consistent with DAF-7/TGFβ and DAF-11 pathways functioning in parallel (Thomas et al., 1993), DAF-11 likely also functions upstream of TGFβ-like and insulin-like signaling. Ohshima and colleagues isolated a daf-11 allele in a genetic screen for mutants exhibiting enhanced dauer arrest and reduced daf-7::GFP expression. Specific expression of DAF-11 in the ASI neuron rescued both dauer arrest as well as daf-7::GFP expression, indicating that DAF-11 functions cell autonomously to promote daf-7 transcription (Murakami et al., 2001). Intriguingly, the insulin-like molecule DAF-28 is also expressed in ASI, and expression of a daf-28::GFP promoter fusion is significantly reduced in daf-11 mutants (Li et al., 2003). Thus, DAF-11 may promote reproductive growth at least in part by promoting cell-autonomous expression of insulin-like and TGFβ-like ligands in sensory neurons.
The Daf-c gene daf-9 and the Daf-d gene daf-12 function downstream of most other proteins that regulate dauer arrest (Albert and Riddle, 1988; Gerisch et al., 2001; Jia et al., 2002; Riddle et al., 1981; Thomas et al., 1993). daf-9 and daf-12 mutants have similar pleiotropies in gonadal migration that are unique among Daf mutants; alleles of both were isolated in a genetic screen for mutants with distal tip cell migration defects (Antebi et al., 1998). Thus, daf-9 and daf-12 define a regulatory module distinct from the guanylyl cyclase, TGFβ-like, and insulin-like pathways.
Multiple lines of evidence are consistent with a model whereby DAF-9 and DAF-12 participate in a steroid hormone pathway that regulates dauer arrest. First, cholesterol deprivation phenocopies daf-9 mutants (Gerisch et al., 2001). Furthermore, DAF-9 functions non-autonomously to inhibit dauer arrest (Gerisch and Antebi, 2004; Mak and Ruvkun, 2004), consistent with its effects being mediated by a hormone. Most strikingly, cloning of daf-9 and daf-12 revealed that daf-9 encodes a cytochrome P450 similar to steroid hydroxylases (Gerisch et al., 2001; Jia et al., 2002), and daf-12 encodes a nuclear receptor homolog with highest similarity in its DNA binding domain to vertebrate pregnane X and vitamin D receptors and in its ligand binding domain to vertebrate thyroid hormone receptors (Antebi et al., 2000). The expanded family of nuclear receptors in C. elegans is described in detail in WormBook (see Nuclear hormone receptors in C. elegans).
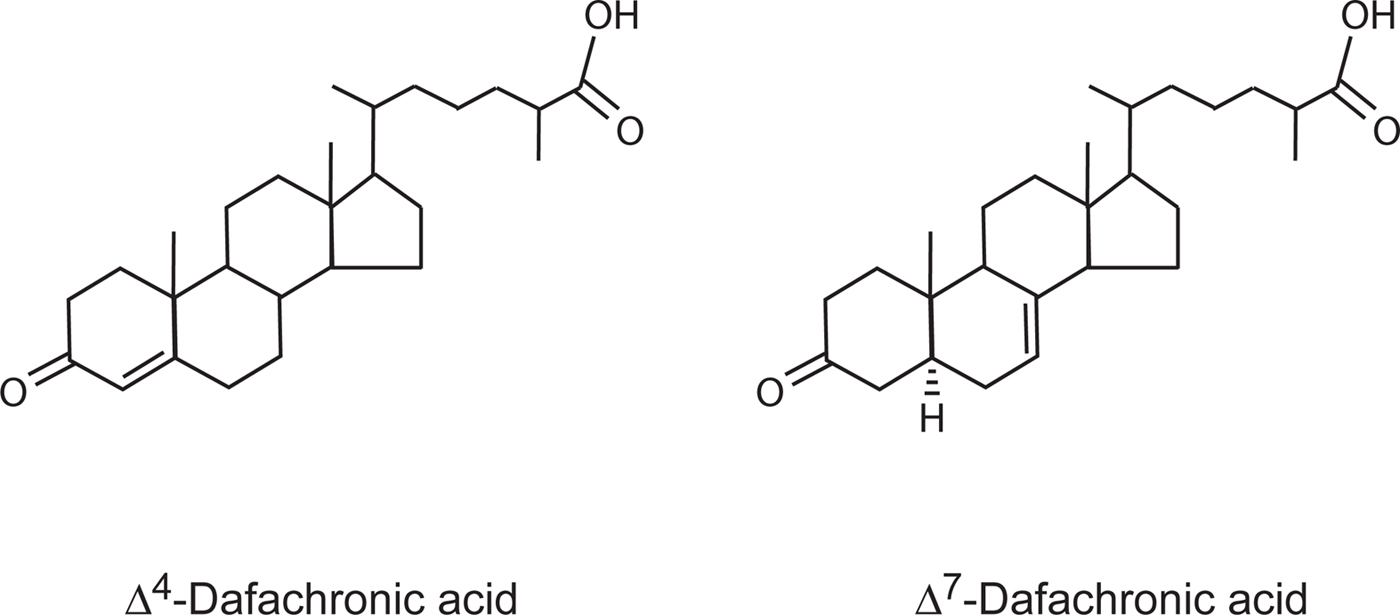
Dauer arrest in daf-9 mutants requires DAF-12 (Gerisch et al., 2001; Jia et al., 2002), indicating that DAF-9 normally inhibits DAF-12 function. Also, daf-12 ligand binding domain mutants have a Daf-c phenotype, suggesting that these mutants are insensitive to DAF-9 inhibition by virtue of their reduced ability to bind ligand (Antebi et al., 2000). Recently, endogenous steroids that are metabolites of DAF-9 and high-affinity ligands for DAF-12 have been identified. DAF-9 is a 3-keto-sterol-26-monooxygenase that synthesizes the DAF-12 ligands Δ4-dafachronic acid and Δ7-dafachronic acid (Motola et al., 2006; see Figure 2).
Daf-c mutants whose phenotypes are exacerbated by cholesterol deprivation may define other components of this pathway (Li et al., 2004; Ohkura et al., 2003; Rottiers et al., 2006). daf-36 encodes a Rieske oxygenase that functions upstream of DAF-9 in dafachronic acid synthesis (Rottiers et al., 2006). Intriguingly, although daf-9 is expressed in the hypodermis, somatic gonad, and XXX cells (Gerisch et al., 2001; Jia et al., 2002; Ohkura et al., 2003), daf-36 is expressed primarily in the intestine and is not present in the XXX cells (Rottiers et al., 2006). It is not clear whether intestinal DAF-36 metabolites are actively transported to DAF-9-expressing tissues or whether they diffuse into neighboring tissues. Mutants of the Niemann-Pick C (NPC) homologs ncr-1 and ncr-2 are weakly Daf-c, but a ncr-2;ncr-1 double mutant has a strong Daf-c phenotype (Li et al., 2004; Sym et al., 2000). The NPC genes encode proteins thought to be involved in intracellular cholesterol trafficking (Chang et al., 2005). NPC1 mutant mice exhibit decreased levels of neurosteroids (Griffin et al., 2004). Analogously, ncr-2;ncr-1 double mutants may undergo dauer arrest as a consequence of reduced dafachronic acid synthesis. Mutations in sdf-9, a gene identified in a synthetic dauer formation screen, cause weak dauer arrest phenotypes that are rescued by overexpression of wild-type daf-9 (Ohkura et al., 2003).
Three mutants initially isolated based on defects in dauer arrest have structural abnormalities in the cilia of chemosensory neurons (see The sensory cilia of C. elegans, and Chemosensation in C. elegans). These ciliary structure abnormalities prevent the amphid neurons from taking up lipophilic dyes, resulting in a Dyf (for abnormal dye filling) phenotype (Perkins et al., 1986; Starich et al., 1995). One of them, daf-10(e1387), was originally isolated as a mutant that did not form dauers in response to starvation (Albert et al., 1981). daf-10 and most other Dyf mutants suppress the dauer arrest phenotype of daf-11 mutants (Starich et al., 1995; Vowels and Thomas, 1992), indicating that cilium structure abnormalities prevent sensory neurons from promoting dauer arrest in the absence of DAF-11 guanylyl cyclase signaling. This may be due in part to excessive activation of insulin-like signaling, as Dyf mutants exhibit enhanced daf-28::GFP expression (Li et al., 2003). daf-10 encodes a protein with several WD repeats that is homologous to IFT122, a component of the intraflagellar transport particle in Chlamydomonas (Bell et al., 2006; Qin et al., 2001).
daf-6(e1377) was also identified based on its inability to form dauers under starvation conditions (Albert et al., 1981). However, in contrast to most Dyf mutants, which have primary defects in sensory neuron ciliary structure (Perkins et al., 1986), dye-filling abnormalities in daf-6 mutants appear to be secondary to defects in the amphid channel that is formed by the glial sheath and socket cells (Albert et al., 1981; Perens and Shaham, 2005; Perkins et al., 1986). These defects prevent the amphid neurons from coming into direct contact with the external environment (Albert et al., 1981; Perens and Shaham, 2005). daf-6 mutants are also unique among Dyf mutants in their inability to suppress the daf-11 dauer arrest phenotype (Vowels and Thomas, 1992), indicating that dauer arrest due to an absence of DAF-11 guanylyl cyclase activity does not require proper formation of amphid channels. The observation that the amphid cilia cannot access the external environment in daf-6 mutants suggests that the Daf-d phenotype of these mutants may be a result of the inability of the amphid neurons to sense dauer pheromone. daf-6 encodes a Patched-related protein expressed in amphid sheath and socket glial cells that is required for lumen formation of the amphid channel (Perens and Shaham, 2005).
All sensory neurons completely lack cilia in daf-19 mutants, the first of which was isolated in Don Riddle's lab (Perkins et al., 1986; Swoboda et al., 2000). daf-19 encodes a RFX-type transcription factor that directly regulates a number of cilia genes via a consensus binding site known as the X box (Swoboda et al., 2000). Mutation of many of these genes individually results in a Dyf phenotype (Swoboda et al., 2000). Unlike all other Dyf mutants, which are Daf-d at 25°C, daf-19 mutants are strongly Daf-c at 25°C (Malone and Thomas, 1994; Swoboda et al., 2000). The mechanisms underlying this observation are likely to be complex and are poorly understood. The severity of the cilia defect in daf-19 mutants, which affects all ciliated neurons, is probably secondary to the misregulation of many genes involved in cilia formation and maintenance and is more widespread than defects in most Dyf mutants. Indeed, laser ablation of all amphid and phasmid neurons results in a Daf-c phenotype (Bargmann and Horvitz, 1991), suggesting that dauer arrest may be a summation phenotype resulting from complete disruption of chemosensory neuron function. Other Dyf mutants may have functional defects in subsets of sensory neurons, some of which may promote dauer arrest and others of which may inhibit dauer arrest. Furthermore, since DAF-19 is the master transcriptional regulator of ciliary genes (Blacque et al., 2005; Swoboda et al., 2000), genes expressed in ciliated neurons whose products may transduce dauer-inhibitory signals (Blacque et al., 2005) may be expressed at levels below the threshold needed to promote reproductive growth in daf-19 mutants. Finally, the DAF-11 guanylyl cyclase that normally promotes reproductive growth is expressed in the cilia of the ASI and ASJ amphid neurons, and mutation of daf-19 could disrupt DAF-11 function, resulting in dauer arrest secondary to low cGMP levels in ASI and ASJ, reduced DAF-7/TGFβ expression in ASI (Murakami et al., 2001), reduced DAF-28/insulin expression in ASI and ASJ (Li et al., 2003), or a combination thereof. Indeed, a kin-8 mutant exhibits dye-filling defects in the ASI amphid neuron and reduced expression of daf-7p::GFP in ASI. However, kin-8 also functions in other cells to regulate dauer arrest, as expression of kin-8 under the control of the daf-7 promoter rescues the ASI dye-filling defect but does not rescue the dauer arrest phenotype of a kin-8 null mutant (Koga et al., 1999). It is not known whether 8-bromo-cGMP can rescue dauer arrest or whether DAF-7::GFP or DAF-28::GFP expression is reduced in daf-19 mutants. Although dauer arrest in daf-19 mutants is suppressed by mutations in daf-16/FoxO, which encodes the major target of DAF-2/insulin-like signaling, daf-19 mutations are epistatic to mutations in daf-3 and daf-5, which encode the main targets of DAF-7/TGFβ-like signaling (Peter Swoboda, personal communication). Thus, reduction of DAF-7/TGFβ expression in ASI cannot fully account for the Daf-c phenotype of daf-19 mutants.
A capricious incubator in Jim Thomas' lab led to the discovery of a plethora of genes whose products function to inhibit dauer arrest at 27°C (Ailion and Thomas, 2000; Ailion and Thomas, 2003). At this temperature, wild-type animals can also arrest as dauers. Therefore, although temperature has a modest influence on dauer arrest at 25°C, at 27°C it can mobilize regulatory molecules and pathways necessary for complete dauer arrest and morphogenesis, albeit at low penetrance (Ailion and Thomas, 2000). The discovery of this phenotype, known as the Hid phenotype (for high temperature induction of dauer), has greatly broadened the repertoire of molecules known to influence dauer arrest (Ailion and Thomas, 2000; Ailion and Thomas, 2003). Many Hid mutants have defects in synaptic transmission (Ailion et al., 1999; Ailion and Thomas, 2000; Mahoney et al., 2006), highlighting the neuroendocrine nature of dauer regulation.
Intriguingly, a subset of Hid mutants that form dauers at high penetrance at 27°C have a Daf-d phenotype at 25°C, including daf-3/SMAD (as described in Section 6.2) and many Dyf mutants (Ailion and Thomas, 2000; Apfeld and Kenyon, 1999). Thus, in Dyf mutants, a mere 2°C elevation in ambient temperature changes the collective dauer-inhibitory output of chemosensory neurons to a dauer-promoting output. The Hid phenotype of Dyf mutants is suppressed by mutations in daf-16 and daf-5 (Ailion and Thomas, 2000; Apfeld and Kenyon, 1999), suggesting that sensory input couples downstream to insulin-like and TGFβ-like pathways.
Laser ablation experiments indicate that temperature elevations may affect the dauer regulatory functions of specific chemosensory neurons. Whereas ablation of both ADF and ASI is necessary to induce dauer arrest in wild-type animals at 20°C (Bargmann and Horvitz, 1991), ablation of ASI alone suffices to induce dauer arrest at 27°C (Ailion and Thomas, 2000). Therefore, high temperature may antagonize the dauer-inhibitory activity of ADF. Alternatively, sensory neurons that do not regulate dauer arrest at lower temperatures may be recruited at 27°C to promote dauer arrest. DAF-7::GFP expression in ASI decreases with increasing temperature (Schackwitz et al., 1996), and it is further decreased at 27°C (Ailion and Thomas, 2000), suggesting a mechanism by which elevated temperature could alter the dauer regulatory output of specific sensory neurons. daf-28::GFP expression is temperature-independent (Li et al., 2003).
Since genetic screens for dauer arrest mutants at 25°C have been saturated, some groups have sought to identify mutants that exhibit dauer arrest phenotypes in specific genetic backgrounds. Similar to the Hid screen (Ailion and Thomas, 2003), such screens allow the identification of genes with relatively subtle influences on dauer arrest.
unc-31 encodes the C. elegans ortholog of CAPS, a cytosolic protein required for calcium-triggered synaptic vesicle release (Ann et al., 1997). unc-31(e169) mutants are Hid (Ailion et al., 1999). In an effort to identify novel genes involved in neuronal function, Isao Katsura's group performed a screen for synthetic dauer formation (sdf) mutants that undergo dauer arrest in an unc-31(e169) mutant background. Since laser ablation of ASI suffices to induce dauer arrest in an unc-31(e169) mutant (Ailion and Thomas, 2000), this screen would potentially identify genes required for ASI specification, development, and function.
Three sdf genes have been described. sdf-13 encodes a TBX2/TBX3 transcription factor homolog that is required for olfactory adaptation mediated by the AWC olfactory neuron (Miyahara et al., 2004), and sdf-14 is allelic to mrp-1, which encodes a multidrug resistance-associated protein that functions in multiple tissues to inhibit dauer arrest (Yabe et al., 2005). Epistasis analysis did not unequivocally reveal the pathway in which mrp-1 acts. sdf-9 encodes a catalytically inactive protein tyrosine phosphatase (PTP)-like molecule that may function in both steroid hormone and insulin-like pathways (Hu et al., 2006; Ohkura et al., 2003). sdf-9 mutants have a weak Daf-c phenotype that is exacerbated by cholesterol deprivation (Ohkura et al., 2003). Strikingly, sdf-9 is expressed specifically in the XXX cells (Hu et al., 2006; Ohkura et al., 2003). Laser ablation of the XXX cells causes a weak dauer arrest phenotype that requires both DAF-12/NR and DAF-16/FoxO in the rest of the animal (Ohkura et al., 2003), indicating that the XXX cells normally inhibit DAF-12 and DAF-16 non-autonomously, perhaps by secreting a hormone. Since DAF-9/CYP27A1 is also expressed in the XXX cells (Ohkura et al., 2003), SDF-9 may function to inhibit dauer arrest by promoting dafachronic acid synthesis in the XXX cells.
Based on the indirect evidence supporting the existence of a signaling pathway that functions in parallel to AGE-1/PI3K (described in Section 6.3), Gary Ruvkun and his colleagues performed a genetic screen to identify mutants that enhance the dauer arrest phenotype of an akt-1 null mutant (eak). 21 independent mutants defining seven complementation groups have been isolated, and three eak genes have been molecularly characterized. eak-4 encodes a novel protein with an N-myristoylation motif, eak-5 is allelic to sdf-9, and eak-6 encodes a PTP-like protein similar to SDF-9 (Hu et al., 2006). Notably, as is the case for sdf-9 (Ohkura et al., 2003), transcriptional and translational GFP fusions of eak-4 and eak-6 are also specifically expressed in the XXX cells (Hu et al., 2006). Translational GFP fusions of all three genes localize to the plasma membrane of the XXX cells, and double mutant combinations among all three genes do not show phenotypic enhancement, suggesting that products of these three genes may function together as a complex at the plasma membrane to promote DAF-9/CYP27A1 activity and steroid hormone synthesis in the XXX cells (Hu et al., 2006).
Although the role of specific amphid neurons in the regulation of dauer arrest has been established for some time (Bargmann and Horvitz, 1991; Schackwitz et al., 1996), how their outputs are integrated during the dauer decision has remained a mystery. The sdf and eak modifier screens have focused attention on the XXX cells as potential sites of integration of insulin-like and steroid hormone signaling.
The dauer arrest phenotype caused by laser ablation of the XXX cells demonstrated their role in preventing dauer arrest and implicated them as endocrine cells (Ohkura et al., 2003). Identification of the XXX cells as daf-9-expressing cells (Ohkura et al., 2003) confirmed their role in the steroid hormone pathway and suggested that they may be sites of dafachronic acid synthesis and secretion.
The ability of both daf-12 and daf-16/FoxO loss-of-function mutations to suppress dauer arrest caused by XXX ablation (Ohkura et al., 2003) was the first indication that the XXX cells couple to both steroid hormone and insulin-like pathways. This was underscored by the discovery that three eak genes are also specifically expressed in the XXX cells and function upstream of daf-12 and daf-16/FoxO (Hu et al., 2006). Specific expression of akt-1 in XXX suppresses the dauer arrest phenotype of eak-4;akt-1 double mutants (Hu et al., 2006), suggesting that insulin-like signaling in XXX can promote dauer bypass, perhaps by potentiating the synthesis and/or secretion of dafachronic acids. Thus, in one model for how insulin-like and steroid hormone signaling are integrated in the XXX cells, insulin-like molecules secreted from amphid neurons (Li et al., 2003; Pierce et al., 2001) could engage DAF-2/InsR on the XXX cell surface, activate AKT-1, and promote dafachronic acid synthesis by DAF-9/CYP27A1.
It remains to be seen whether daf-2/InsR is expressed in the XXX cells, as well as whether DAF-7/TGFβ-like signals converge on the XXX cells. Furthermore, how XXX outputs regulate DAF-16/FoxO nonautonomously (Ohkura et al., 2003) is not understood. One obvious hypothesis is that the XXX cells may secrete insulin-like molecules as well as steroid hormones. It will be of great interest to determine the expression profile of the XXX cells.
It should be noted that, although the XXX cells clearly play a role in regulating dauer development, the relative weakness of the XXX laser ablation dauer arrest phenotype (Ohkura et al., 2003) compared to the dauer arrest phenotype observed in strong daf-2/InsR, daf-7/TGFβ, and steroid hormone pathway mutants indicates that other cells and tissues besides XXX likely play major roles in the dauer decision.
Gene expression profiles have been generated using serial analysis of gene expression (SAGE; Jones et al., 2001) and microarray-based profiling (Wang and Kim, 2003) to identify gene expression changes that correlate with dauer arrest. Marra and colleagues performed SAGE on wild-type dauers generated by starvation and mixed-stage animals (both grown in liquid culture; Jones et al., 2001). This mixed-stage culture was highly enriched in L1 larvae. They identified 358 dauer-specific and 533 mixed-stage-specific transcripts (P = 0.05). The most abundant dauer transcript was the non-coding transcript tts-1. tts-1 was enriched 20-fold in dauers compared to mixed-stage animals. The most highly expressed dauer-specific transcript corresponded to the small heat shock protein hsp-12.6. Enrichment of the variant histone H1 hil-1 and the nucleosome assembly protein D2096.8 in dauers suggests that dauer chromatin structure may differ from that of growing animals (Jones et al., 2001).
To obtain a picture of the dynamics of gene expression accompanying exit from dauer arrest, Wang and Kim performed a 12-hour time course of microarray-based expression profiles on dauers after recovery was triggered by refeeding on plates (Wang and Kim, 2003). To control for gene expression changes induced by refeeding per se, they compared these profiles to those generated after refeeding of starved L1 larvae as well as to a mixed-stage sample. They clustered 2430 genes that change expression during dauer exit (P ≤ 0.001) into five groups based on time course kinetics.
Although genes such as hsp-12.6 and sod-3 were identified as dauer-enriched transcripts by both SAGE and microarray-based profiling, there was strikingly little overlap in the expression profiles generated using the two different approaches (Jones et al., 2001; Wang and Kim, 2003). The biological significance of these expression changes remains to be determined.
Using the same platform as Wang and Kim (Wang and Kim, 2003), Garth Patterson and his colleagues compared gene expression profiles of daf-7/TGFβ pathway mutants undergoing dauer arrest to profiles of wild-type early L3 larvae (Liu et al., 2004). They identified over 1200 genes whose expression was regulated strongly in the daf-7/TGFβ pathway mutants undergoing dauer arrest (P = 0.01). Notably, daf-2/InsR and daf-9 are downregulated and daf-12 is upregulated in daf-7/TGFβ pathway mutants, suggesting that DAF-7/TGFβ signaling feeds forward to promote insulin-like and steroid hormone signaling during larval development in replete environments. daf-9 expression increases early and remains elevated during dauer exit (Wang and Kim, 2003), suggesting that modulation of steroid hormone signaling is an important aspect of dauer recovery as well as dauer bypass in replete environments.
In an effort to obtain a more panoramic view of the DAF-7/TGFβ signaling landscape (see TGF-β signaling), Marc Vidal, Gary Ruvkun, and their colleagues combined interactome mapping with RNAi-based genetic analysis to define a dauer regulatory network (Tewari et al., 2004). They performed two rounds of yeast two-hybrid screening starting with all known components of DAF-7/TGFβ signaling as baits, and they tested interacting proteins for roles in dauer regulation using dauer formation assays after gene-specific RNAi in wild-type and various TGFβ pathway mutant backgrounds. The approach was validated by their identification of daf-5, a Sno oncogene homolog that is a major target of DAF-7/TGFβ signaling and that was cloned independently using classical forward genetic analysis (Da Graca et al., 2004). This strategy yielded eight candidate modulators of DAF-7/TGFβ signaling that had not been identified in classical genetic screens for pathway components.
Genome-wide RNAi screens for dauer arrest genes have not been reported, perhaps because RNAi of known dauer-constitutive genes does not efficiently phenocopy mutants (Tewari et al., 2004). In light of the success of modifier screens in identifying new components of dauer regulatory pathways (see Section 9), RNAi-based enhancer screens may turn out to be a fruitful strategy to identify new dauer genes.
Over the past decade, much has been learned about the molecular and cellular underpinnings of the regulation of dauer arrest. In particular, significant progress has been made in defining the components of signal transduction pathways that regulate dauer arrest. However, many outstanding questions remain in the field. More than twenty years ago, the fundamental influences on dauer arrest were defined (Golden and Riddle, 1982; Golden and Riddle, 1984a; Golden and Riddle, 1984b). Remarkably, the mechanistic basis for the effect of pheromone, food, and temperature on dauer arrest has resisted elucidation. The availability of synthetic dauer pheromone components (Jeong et al., 2005; Butcher et al., 2007) should facilitate the search for specific pheromone receptors. Other critical questions that remain to be answered include the molecular identity of the food signal, how it is interpreted by the animal, and how changes in ambient temperature are translated into molecular events that influence dauer arrest. It is likely that these environmental signals will function primarily by modulating signaling flux through the guanylyl cyclase, TGFβ-like, insulin-like, and hormonal pathways described here.
Sequencing of the C. elegans genome has identified three expanded families of molecules that play critical roles in regulating dauer arrest. The C. elegans genome contains 40 genes encoding insulin-like molecules, 83 genes encoding cytochromes P450, and 284 genes encoding nuclear receptors. Unequivocal roles in dauer biology have been established for exactly one molecule from each family (the insulin-like molecule DAF-28 (Li et al., 2003), the CYP27A1 functional ortholog DAF-9 (Gerisch et al., 2001; Jia et al., 2002; Motola et al., 2006)), and the nuclear receptor DAF-12 (Antebi et al., 2000; Motola et al., 2006). It is likely that future research will unveil roles for other members of these families in regulating dauer arrest.
Finally, the interface among signal transduction pathways that regulate dauer arrest will continue to be an active area of investigation in the near future. Although epistasis analysis has contributed substantially to our understanding of how these signaling pathways interact, further studies are certain to yield greater insight into the complexities of their interdigitations. These studies are relevant not only to dauer biology but also to human health, as the evolutionary conservation of these signal transduction pathways suggests that what we learn about their interactions during C. elegans larval development may be germane to the interactions of similar signaling pathways in the pathogenesis of common diseases such as diabetes mellitus and cancer.
I thank Peter Swoboda and Weiqing Li for providing unpublished results and Rebecca Butcher and Jon Clardy for providing data prior to publication. Research in my laboratory is funded by the National Institute of Diabetes and Digestive and Kidney Diseases and the Michigan Diabetes Research and Training Center at the University of Michigan Medical School.
Ailion, M., Inoue, T., Weaver, C.I., Holdcraft, R.W., and Thomas, J.H. (1999). Neurosecretory control of aging in Caenorhabditis elegans. Proc. Natl. Acad. Sci. U.S.A. 96, 7394–7397. Abstract Article
Ailion, M., and Thomas, J.H. (2000). Dauer formation induced by high temperatures in Caenorhabditis elegans. Genetics 156, 1047–1067. Abstract
Ailion, M., and Thomas, J.H. (2003). Isolation and characterization of high-temperature-induced dauer formation mutants in Caenorhabditis elegans. Genetics 165, 127–144. Abstract
Albert, P.S., Brown, S.J., and Riddle, D.L. (1981). Sensory control of dauer larva formation in Caenorhabditis elegans. J. Comp. Neurol. 198, 435–451. Abstract Article
Albert, P.S., and Riddle, D.L. (1988). Mutants of Caenorhabditis elegans that form dauer-like larvae. Dev. Biol. 126, 270–293. Abstract Article
Ann, K., Kowalchyk, J.A., Loyet, K.M., and Martin, T.F. (1997). Novel Ca2+-binding protein (CAPS) related to UNC-31 required for Ca2+-activated exocytosis. J. Biol. Chem. 272, 19637–19640. Abstract Article
Antebi, A., Culotti, J.G., and Hedgecock, E.M. (1998). daf-12 regulates developmental age and the dauer alternative in Caenorhabditis elegans. Development 125, 1191–1205. Abstract
Antebi, A., Yeh, W.H., Tait, D., Hedgecock, E.M., and Riddle, D.L. (2000). daf-12 encodes a nuclear receptor that regulates the dauer diapause and developmental age in C. elegans. Genes Dev. 14, 1512–1527. Abstract
Apfeld, J., and Kenyon, C. (1998). Cell nonautonomy of C. elegans daf-2 function in the regulation of diapause and life span. Cell 95, 199–210. Abstract Article
Apfeld, J., and Kenyon, C. (1999). Regulation of lifespan by sensory perception in Caenorhabditis elegans. Nature 402, 804–809. Abstract Article
Bargmann, C.I., and Horvitz, H.R. (1991). Control of larval development by chemosensory neurons in Caenorhabditis elegans. Science 251, 1243–1246. Abstract
Bell, L.R., Stone, S., Yochem, J., Shaw, J.E., and Herman, R.K. (2006). The molecular identities of the Caenorhabditis elegans intraflagellar transport genes dyf-6, daf-10 and osm-1. Genetics 173, 1275–1286. Abstract Article
Berdichevsky, A., Viswanathan, M., Horvitz, H.R., and Guarente, L. (2006). C. elegans SIR-2.1 interacts with 14-3-3 proteins to activate DAF-16 and extend life span. Cell 125, 1165–1177. Abstract Article
Birnby, D.A., Link, E.M., Vowels, J.J., Tian, H., Colacurcio, P.L., and Thomas, J.H. (2000). A transmembrane guanylyl cyclase (DAF-11) and Hsp90 (DAF-21) regulate a common set of chemosensory behaviors in Caenorhabditis elegans. Genetics 155, 85–104. Abstract
Blacque, O.E., Perens, E.A., Boroevich, K.A., Inglis, P.N., Li, C., Warner, A., Khattra, J., Holt, R.A., Ou, G., Mah, A.K., et al. (2005). Functional genomics of the cilium, a sensory organelle. Curr. Biol. 15, 935–941. Abstract Article
Butcher, R.A., Fujita, M., Schroeder, F.C., and Clardy, J. (2007). Small-molecule pheromones that control dauer development in Caenorhabditis elegans. Nat. Chem. Biol. 3, 420–422. Abstract Article
Cassada, R.C., and Russell, R.L. (1975). The dauerlarva, a post-embryonic developmental variant of the nematode Caenorhabditis elegans. Dev. Biol. 46, 326–342. Abstract Article
Chang, T.Y., Reid, P.C., Sugii, S., Ohgami, N., Cruz, J.C., and Chang, C.C. (2005). Niemann-Pick type C disease and intracellular cholesterol trafficking. J. Biol. Chem. 280, 20917–20920. Abstract Article
Coburn, C.M., and Bargmann, C.I. (1996). A putative cyclic nucleotide-gated channel is required for sensory development and function in C. elegans. Neuron 17, 695–706. Abstract Article
Coburn, C.M., Mori, I., Ohshima, Y., and Bargmann, C.I. (1998). A cyclic nucleotide-gated channel inhibits sensory axon outgrowth in larval and adult Caenorhabditis elegans: a distinct pathway for maintenance of sensory axon structure. Development 125, 249–258. Abstract
Da Graca, L.S., Zimmerman, K.K., Mitchell, M.C., Kozhan-Gorodetska, M., Sekiewicz, K., Morales, Y., and Patterson, G.I. (2004). DAF-5 is a Ski oncoprotein homolog that functions in a neuronal TGF-beta pathway to regulate C. elegans dauer development. Development 131, 435–446. Abstract Article
Dalley, B.K., and Golomb, M. (1992). Gene expression in the Caenorhabditis elegans dauer larva: developmental regulation of Hsp90 and other genes. Dev. Biol. 151, 80–90. Abstract Article
Daniels, S.A., Ailion, M., Thomas, J.H., and Sengupta, P. (2000). egl-4 acts through a transforming growth factor-beta/SMAD pathway in Caenorhabditis elegans to regulate multiple neuronal circuits in response to sensory cues. Genetics 156, 123–141. Abstract
Dillin, A., Crawford, D.K., and Kenyon, C. (2002). Timing requirements for insulin/IGF-1 signaling in C. elegans. Science 298, 830–834. Abstract Article
Estevez, A.O. (1997). The role of the daf-8 gene in Caenorhabditis elegans dauer larva development, University of Missouri, Columbia, Columbia.
Estevez, M., Attisano, L., Wrana, J.L., Albert, P.S., Massague, J., and Riddle, D.L. (1993). The daf-4 gene encodes a bone morphogenetic protein receptor controlling C. elegans dauer larva development. Nature 365, 644–649. Abstract Article
Friedman, D.B., and Johnson, T.E. (1988). A mutation in the age-1 gene in Caenorhabditis elegans lengthens life and reduces hermaphrodite fertility. Genetics 118, 75–86. Abstract
Fujiwara, M., Sengupta, P., and McIntire, S.L. (2002). Regulation of body size and behavioral state of C. elegans by sensory perception and the EGL-4 cGMP-dependent protein kinase. Neuron 36, 1091–1102. Abstract Article
Gems, D., Sutton, A.J., Sundermeyer, M.L., Albert, P.S., King, K.V., Edgley, M.L., Larsen, P.L., and Riddle, D.L. (1998). Two pleiotropic classes of daf-2 mutation affect larval arrest, adult behavior, reproduction and longevity in Caenorhabditis elegans. Genetics 150, 129–155. Abstract
Georgi, L.L., Albert, P.S., and Riddle, D.L. (1990). daf-1, a C. elegans gene controlling dauer larva development, encodes a novel receptor protein kinase. Cell 61, 635–645. Abstract Article
Gerisch, B., and Antebi, A. (2004). Hormonal signals produced by DAF-9/cytochrome P450 regulate C. elegans dauer diapause in response to environmental cues. Development 131, 1765–1776. Abstract Article
Gerisch, B., Weitzel, C., Kober-Eisermann, C., Rottiers, V., and Antebi, A. (2001). A hormonal signaling pathway influencing C. elegans metabolism, reproductive development, and life span. Dev. Cell 1, 841–851. Abstract Article
Gil, E.B., Malone Link, E., Liu, L.X., Johnson, C.D., and Lees, J.A. (1999). Regulation of the insulin-like developmental pathway of Caenorhabditis elegans by a homolog of the PTEN tumor suppressor gene. Proc. Natl. Acad. Sci. U.S.A. 96, 2925–2930. Abstract Article
Golden, J.W., and Riddle, D.L. (1982). A pheromone influences larval development in the nematode Caenorhabditis elegans. Science 218, 578–580. Abstract
Golden, J.W., and Riddle, D.L. (1984a). The Caenorhabditis elegans dauer larva: developmental effects of pheromone, food, and temperature. Dev. Biol. 102, 368–378. Abstract Article
Golden, J.W., and Riddle, D.L. (1984b). A pheromone-induced developmental switch in Caenorhabditis elegans: Temperature-sensitive mutants reveal a wild-type temperature-dependent process. Proc. Natl. Acad. Sci. U.S.A. 81, 819–823. Abstract Article
Golden, J.W., and Riddle, D.L. (1985). A gene affecting production of the Caenorhabditis elegans dauer-inducing pheromone. Mol. Gen. Genet. 198, 534–536. Abstract Article
Gottlieb, S., and Ruvkun, G. (1994). daf-2, daf-16 and daf-23: genetically interacting genes controlling Dauer formation in Caenorhabditis elegans. Genetics 137, 107–120. Abstract
Griffin, L.D., Gong, W., Verot, L., and Mellon, S.H. (2004). Niemann-Pick type C disease involves disrupted neurosteroidogenesis and responds to allopregnanolone. Nat. Med. 10, 704–711. Abstract Article
Gunther, C.V., Georgi, L.L., and Riddle, D.L. (2000). A Caenorhabditis elegans type I TGF beta receptor can function in the absence of type II kinase to promote larval development. Development 127, 3337–3347. Abstract
Hertweck, M., Gobel, C., and Baumeister, R. (2004). C. elegans SGK-1 is the critical component in the Akt/PKB kinase complex to control stress response and life span. Dev. Cell 6, 577–588. Abstract Article
Hu, P.J., Xu, J., and Ruvkun, G. (2006). Two membrane-associated tyrosine phosphatase homologs potentiate C. elegans AKT-1/PKB signaling. PLoS Genet. 2, e99. Abstract Article
Hua, Q.X., Nakagawa, S.H., Wilken, J., Ramos, R.R., Jia, W., Bass, J., and Weiss, M.A. (2003). A divergent INS protein in Caenorhabditis elegans structurally resembles human insulin and activates the human insulin receptor. Genes Dev. 17, 826–831. Abstract Article
Inoki, K., and Guan, K.L. (2006). Complexity of the TOR signaling network. Trends Cell Biol. 16, 206–212. Abstract Article
Inoue, T., and Thomas, J.H. (2000a). Suppressors of transforming growth factor-beta pathway mutants in the Caenorhabditis elegans dauer formation pathway. Genetics 156, 1035–1046. Abstract
Inoue, T., and Thomas, J.H. (2000b). Targets of TGF-beta signaling in Caenorhabditis elegans dauer formation. Dev. Biol. 217, 192–204. Abstract Article
Jeong, P.Y., Jung, M., Yim, Y.H., Kim, H., Park, M., Hong, E., Lee, W., Kim, Y.H., Kim, K., and Paik, Y.K. (2005). Chemical structure and biological activity of the Caenorhabditis elegans dauer-inducing pheromone. Nature 433, 541–545. Abstract Article
Jia, K., Albert, P.S., and Riddle, D.L. (2002). DAF-9, a cytochrome P450 regulating C. elegans larval development and adult longevity. Development 129, 221–231. Abstract
Jia, K., Chen, D., and Riddle, D.L. (2004). The TOR pathway interacts with the insulin signaling pathway to regulate C. elegans larval development, metabolism and life span. Development 131, 3897–3906. Abstract Article
Jones, S.J., Riddle, D.L., Pouzyrev, A.T., Velculescu, V.E., Hillier, L., Eddy, S.R., Stricklin, S.L., Baillie, D.L., Waterston, R., and Marra, M.A. (2001). Changes in gene expression associated with developmental arrest and longevity in Caenorhabditis elegans. Genome Res. 11, 1346–1352. Abstract Article
Kao, G., Nordenson, C., Still, M., Ronnlund, A., Tuck, S., and Naredi, P. (2007). ASNA-1 Positively Regulates Insulin Secretion in C. elegans and Mammalian Cells. Cell 128, 577–587. Abstract Article
Kenyon, C., Chang, J., Gensch, E., Rudner, A., and Tabtiang, R. (1993). A C. elegans mutant that lives twice as long as wild type. Nature 366, 461–464. Abstract Article
Kimura, K.D., Tissenbaum, H.A., Liu, Y., and Ruvkun, G. (1997). daf-2, an insulin receptor-like gene that regulates longevity and diapause in Caenorhabditis elegans. Science 277, 942–946. Abstract Article
Klass, M., and Hirsh, D. (1976). Non-ageing developmental variant of Caenorhabditis elegans. Nature 260, 523–525. Abstract Article
Koga, M., Take-uchi, M., Tameishi, T., and Ohshima, Y. (1999). Control of DAF-7 TGF-(alpha) expression and neuronal process development by a receptor tyrosine kinase KIN-8 in Caenorhabditis elegans. Development 126, 5387–5398. Abstract
Komatsu, H., Mori, I., Rhee, J.S., Akaike, N., and Ohshima, Y. (1996). Mutations in a cyclic nucleotide-gated channel lead to abnormal thermosensation and chemosensation in C. elegans. Neuron 17, 707–718. Abstract Article
L'Etoile, N.D., Coburn, C.M., Eastham, J., Kistler, A., Gallegos, G., and Bargmann, C.I. (2002). The cyclic GMP-dependent protein kinase EGL-4 regulates olfactory adaptation in C. elegans. Neuron 36, 1079–1089. Abstract Article
Larsen, P.L. (1993). Aging and resistance to oxidative damage in Caenorhabditis elegans. Proc. Natl. Acad. Sci. U.S.A. 90, 8905–8909. Abstract Article
Lee, R.Y., Hench, J., and Ruvkun, G. (2001). Regulation of C. elegans DAF-16 and its human ortholog FKHRL1 by the daf-2 insulin-like signaling pathway. Curr. Biol. 11, 1950–1957. Abstract Article
Li, J., Brown, G., Ailion, M., Lee, S., and Thomas, J.H. (2004). NCR-1 and NCR-2, the C. elegans homologs of the human Niemann-Pick type C1 disease protein, function upstream of DAF-9 in the dauer formation pathways. Development 131, 5741–5752. Abstract Article
Li, J., Tewari, M., Vidal, M., and Lee, S.S. (2007). The 14-3-3 protein FTT-2 regulates DAF-16 in Caenorhabditis elegans. Dev. Biol. 301, 82–91. Abstract Article
Li, W., Gao, B., Lee, S.M., Bennett, K., and Fang, D. (2007). RLE-1, an E3 Ubiquitin Ligase, Regulates C. elegans Aging by Catalyzing DAF-16 Polyubiquitination. Dev. Cell 12, 235–246. Abstract Article
Li, W., Kennedy, S.G., and Ruvkun, G. (2003). daf-28 encodes a C. elegans insulin superfamily member that is regulated by environmental cues and acts in the DAF-2 signaling pathway. Genes Dev. 17, 844–858. Abstract Article
Liao, E.H., Hung, W., Abrams, B., and Zhen, M. (2004). An SCF-like ubiquitin ligase complex that controls presynaptic differentiation. Nature 430, 345–350. Abstract Article
Libina, N., Berman, J.R., and Kenyon, C. (2003). Tissue-specific activities of C. elegans DAF-16 in the regulation of lifespan. Cell 115, 489–502. Abstract Article
Lin, K., Dorman, J.B., Rodan, A., and Kenyon, C. (1997). daf-16: An HNF-3/forkhead family member that can function to double the life-span of Caenorhabditis elegans. Science 278, 1319–1322. Abstract Article
Lin, K., Hsin, H., Libina, N., and Kenyon, C. (2001). Regulation of the Caenorhabditis elegans longevity protein DAF-16 by insulin/IGF-1 and germline signaling. Nat. Genet. 28, 139–145. Abstract Article
Liu, T., Zimmerman, K.K., and Patterson, G.I. (2004). Regulation of signaling genes by TGFbeta during entry into dauer diapause in C. elegans. BMC Dev. Biol. 4, 11. Abstract Article
Long, X., Spycher, C., Han, Z.S., Rose, A.M., Muller, F., and Avruch, J. (2002). TOR deficiency in C. elegans causes developmental arrest and intestinal atrophy by inhibition of mRNA translation. Curr. Biol. 12, 1448–1461. Abstract Article
Mahoney, T.R., Liu, Q., Itoh, T., Luo, S., Hadwiger, G., Vincent, R., Wang, Z.W., Fukuda, M., and Nonet, M.L. (2006). Regulation of synaptic transmission by RAB-3 and RAB-27 in Caenorhabditis elegans. Mol. Biol. Cell 17, 2617–2625. Abstract Article
Mak, H.Y., and Ruvkun, G. (2004). Intercellular signaling of reproductive development by the C. elegans DAF-9 cytochrome P450. Development 131, 1777–1786. Abstract Article
Malone, E.A., and Thomas, J.H. (1994). A screen for nonconditional dauer-constitutive mutations in Caenorhabditis elegans. Genetics 136, 879–886. Abstract
Mihaylova, V.T., Borland, C.Z., Manjarrez, L., Stern, M.J., and Sun, H. (1999). The PTEN tumor suppressor homolog in Caenorhabditis elegans regulates longevity and dauer formation in an insulin receptor-like signaling pathway. Proc. Natl. Acad. Sci. U.S.A. 96, 7427–7432. Abstract Article
Miyahara, K., Suzuki, N., Ishihara, T., Tsuchiya, E., and Katsura, I. (2004). TBX2/TBX3 transcriptional factor homologue controls olfactory adaptation in Caenorhabditis elegans. J. Neurobiol. 58, 392–402. Abstract Article
Morita, K., Shimizu, M., Shibuya, H., and Ueno, N. (2001). A DAF-1-binding protein BRA-1 is a negative regulator of DAF-7 TGF-beta signaling. Proc. Natl. Acad. Sci. U.S.A. 98, 6284–6288. Abstract Article
Morris, J.Z., Tissenbaum, H.A., and Ruvkun, G. (1996). A phosphatidylinositol-3-OH kinase family member regulating longevity and diapause in Caenorhabditis elegans. Nature 382, 536–539. Abstract Article
Motola, D.L., Cummins, C.L., Rottiers, V., Sharma, K.K., Li, T., Li, Y., Suino-Powell, K., Xu, H.E., Auchus, R.J., Antebi, A., and Mangelsdorf, D.J. (2006). Identification of ligands for DAF-12 that govern dauer formation and reproduction in C. elegans. Cell 124, 1209–1223. Abstract Article
Murakami, M., Koga, M., and Ohshima, Y. (2001). DAF-7/TGF-beta expression required for the normal larval development in C. elegans is controlled by a presumed guanylyl cyclase DAF-11. Mech. Dev. 109, 27–35. Abstract Article
Murphy, C.T., McCarroll, S.A., Bargmann, C.I., Fraser, A., Kamath, R.S., Ahringer, J., Li, H., and Kenyon, C. (2003). Genes that act downstream of DAF-16 to influence the lifespan of Caenorhabditis elegans. Nature 424, 277–283. Abstract Article
Ogg, S., Paradis, S., Gottlieb, S., Patterson, G.I., Lee, L., Tissenbaum, H.A., and Ruvkun, G. (1997). The Fork head transcription factor DAF-16 transduces insulin-like metabolic and longevity signals in C. elegans. Nature 389, 994–999. Abstract Article
Ogg, S., and Ruvkun, G. (1998). The C. elegans PTEN homolog, DAF-18, acts in the insulin receptor-like metabolic signaling pathway. Mol. Cell 2, 887–893. Abstract Article
Ohkura, K., Suzuki, N., Ishihara, T., and Katsura, I. (2003). SDF-9, a protein tyrosine phosphatase-like molecule, regulates the L3/dauer developmental decision through hormonal signaling in C. elegans. Development 130, 3237–3248. Abstract Article
Oldham, S., and Hafen, E. (2003). Insulin/IGF and target of rapamycin signaling: a TOR de force in growth control. Trends Cell Biol. 13, 79–85. Abstract Article
Paradis, S., Ailion, M., Toker, A., Thomas, J.H., and Ruvkun, G. (1999). A PDK1 homolog is necessary and sufficient to transduce AGE-1 PI3 kinase signals that regulate diapause in Caenorhabditis elegans. Genes Dev. 13, 1438–1452. Abstract
Paradis, S., and Ruvkun, G. (1998). Caenorhabditis elegans Akt/PKB transduces insulin receptor-like signals from AGE-1 PI3 kinase to the DAF-16 transcription factor. Genes Dev. 12, 2488–2498. Abstract
Patterson, G.I., Koweek, A., Wong, A., Liu, Y., and Ruvkun, G. (1997). The DAF-3 Smad protein antagonizes TGF-beta-related receptor signaling in the Caenorhabditis elegans dauer pathway. Genes Dev. 11, 2679–2690. Abstract
Patterson, G.I., and Padgett, R.W. (2000). TGF beta-related pathways. Roles in Caenorhabditis elegans development. Trends Genet. 16, 27–33. Abstract Article
Perens, E.A., and Shaham, S. (2005). C. elegans daf-6 encodes a patched-related protein required for lumen formation. Dev. Cell 8, 893–906. Abstract Article
Perkins, L.A., Hedgecock, E.M., Thomson, J.N., and Culotti, J.G. (1986). Mutant sensory cilia in the nematode Caenorhabditis elegans. Dev. Biol. 117, 456–487. Abstract Article
Pierce, S.B., Costa, M., Wisotzkey, R., Devadhar, S., Homburger, S.A., Buchman, A.R., Ferguson, K.C., Heller, J., Platt, D.M., Pasquinelli, A.A., et al. (2001). Regulation of DAF-2 receptor signaling by human insulin and ins-1, a member of the unusually large and diverse C. elegans insulin gene family. Genes Dev. 15, 672–686. Abstract Article
Qin, H., Rosenbaum, J.L., and Barr, M.M. (2001). An autosomal recessive polycystic kidney disease gene homolog is involved in intraflagellar transport in C. elegans ciliated sensory neurons. Curr. Biol. 11, 457–461. Abstract Article
Ren, P., Lim, C.S., Johnsen, R., Albert, P.S., Pilgrim, D., and Riddle, D.L. (1996). Control of C. elegans larval development by neuronal expression of a TGF-beta homolog. Science 274, 1389–1391. Abstract Article
Riddle, D.L., and Albert, P.S. (1997). Genetic and Environmental Regulation of Dauer Larva Development. In C. elegans II, D.L. Riddle, T. Blumenthal, B.J. Meyer, and J.R. Priess, eds. (Plainview, New York: Cold Spring Harbor Laboratory Press), pp. 739–768.
Riddle, D.L., Swanson, M.M., and Albert, P.S. (1981). Interacting genes in nematode dauer larva formation. Nature 290, 668–671. Abstract Article
Rottiers, V., Motola, D.L., Gerisch, B., Cummins, C.L., Nishiwaki, K., Mangelsdorf, D.J., and Antebi, A. (2006). Hormonal control of C. elegans dauer formation and life span by a Rieske-like oxygenase. Dev. Cell 10, 473–482. Abstract Article
Rouault, J.P., Kuwabara, P.E., Sinilnikova, O.M., Duret, L., Thierry-Mieg, D., and Billaud, M. (1999). Regulation of dauer larva development in Caenorhabditis elegans by daf-18, a homologue of the tumour suppressor PTEN. Curr. Biol. 9, 329–332. Abstract Article
Schackwitz, W.S., Inoue, T., and Thomas, J.H. (1996). Chemosensory neurons function in parallel to mediate a pheromone response in C. elegans. Neuron 17, 719–728. Abstract Article
Shi, Y., and Massague, J. (2003). Mechanisms of TGF-beta signaling from cell membrane to the nucleus. Cell 113, 685–700. Abstract Article
Snutch, T.P., and Baillie, D.L. (1983). Alterations in the pattern of gene expression following heat shock in the nematode Caenorhabditis elegans. Can. J. Biochem. Cell Biol. 61, 480–487. Abstract
Stansberry, J., Baude, E.J., Taylor, M.K., Chen, P.J., Jin, S.W., Ellis, R.E., and Uhler, M.D. (2001). A cGMP-dependent protein kinase is implicated in wild-type motility in C. elegans. J. Neurochem. 76, 1177–1187. Abstract Article
Starich, T.A., Herman, R.K., Kari, C.K., Yeh, W.H., Schackwitz, W.S., Schuyler, M.W., Collet, J., Thomas, J.H., and Riddle, D.L. (1995). Mutations affecting the chemosensory neurons of Caenorhabditis elegans. Genetics 139, 171–188. Abstract
Swoboda, P., Adler, H.T., and Thomas, J.H. (2000). The RFX-type transcription factor DAF-19 regulates sensory neuron cilium formation in C. elegans. Mol. Cell 5, 411–421. Abstract Article
Sym, M., Basson, M., and Johnson, C. (2000). A model for niemann-pick type C disease in the nematode Caenorhabditis elegans. Curr. Biol. 10, 527–530. Abstract Article
Tewari, M., Hu, P.J., Ahn, J.S., Ayivi-Guedehoussou, N., Vidalain, P.O., Li, S., Milstein, S., Armstrong, C.M., Boxem, M., Butler, M.D., et al. (2004). Systematic interactome mapping and genetic perturbation analysis of a C. elegans TGF-beta signaling network. Mol. Cell 13, 469–482. Abstract Article
Thomas, J.H., Birnby, D.A., and Vowels, J.J. (1993). Evidence for parallel processing of sensory information controlling dauer formation in Caenorhabditis elegans. Genetics 134, 1105–1117. Abstract
Vanfleteren, J.R., and De Vreese, A. (1995). The gerontogenes age-1 and daf-2 determine metabolic rate potential in aging Caenorhabditis elegans. FASEB J. 9, 1355–1361. Abstract
Vowels, J.J., and Thomas, J.H. (1992). Genetic analysis of chemosensory control of dauer formation in Caenorhabditis elegans. Genetics 130, 105–123. Abstract
Vowels, J.J., and Thomas, J.H. (1994). Multiple chemosensory defects in daf-11 and daf-21 mutants of Caenorhabditis elegans. Genetics 138, 303–316. Abstract
Wadsworth, W.G., and Riddle, D.L. (1989). Developmental regulation of energy metabolism in Caenorhabditis elegans. Dev. Biol. 132, 167–173. Abstract Article
Wang, J., and Kim, S.K. (2003). Global analysis of dauer gene expression in Caenorhabditis elegans. Development 130, 1621–1634. Abstract Article
Wolkow, C.A., Kimura, K.D., Lee, M.S., and Ruvkun, G. (2000). Regulation of C. elegans life-span by insulinlike signaling in the nervous system. Science 290, 147–150. Abstract Article
Wolkow, C.A., Munoz, M.J., Riddle, D.L., and Ruvkun, G. (2002). Insulin receptor substrate and p55 orthologous adaptor proteins function in the Caenorhabditis elegans daf-2/insulin-like signaling pathway. J. Biol. Chem. 277, 49591–49597. Abstract Article
Yabe, T., Suzuki, N., Furukawa, T., Ishihara, T., and Katsura, I. (2005). Multidrug resistance-associated protein MRP-1 regulates dauer diapause by its export activity in Caenorhabditis elegans. Development 132, 3197–3207. Abstract Article
Yu, S., Avery, L., Baude, E., and Garbers, D.L. (1997). Guanylyl cyclase expression in specific sensory neurons: a new family of chemosensory receptors. Proc. Natl. Acad. Sci. U.S.A. 94, 3384–3387. Abstract Article
Zwaal, R.R., Mendel, J.E., Sternberg, P.W., and Plasterk, R.H. (1997). Two neuronal G proteins are involved in chemosensation of the Caenorhabditis elegans Dauer-inducing pheromone. Genetics 145, 715–727. Abstract
*Edited by Donald L. Riddle. Last Revised May 21, 2007. Published August 08, 2007. This chapter should be cited as: Hu, P.J. Dauer (August 08, 2007), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.144.1, http://www.wormbook.org.
Copyright: © 2007 Patrick J. Hu. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
§To whom correspondence should be addressed. Phone: (734) 615-9656; Fax: (734) 615-5493. E-mail: pathu@umich.edu
 All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.
All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.