Embryo series courtesy of Einhard Schierenberg
Embryo series courtesy of Einhard SchierenbergAbstract
Genetic mosaics can be used to gain insight into the cell specificity of gene function. How Caenorhabditis elegans mosaics are typically generated is reviewed, and several examples with relevance to developmental studies are mentioned. One example is mpk-1, which encodes a member of the Ras-MAP-kinase pathway. mpk-1 mosaics have been a means of studying the distinct cells that require the gene for distinct fates during development. The gene bre-5 is used as an example of the usefulness of mosaic analysis for non-developmental studies. Potential problems with mosaic analysis are discussed, and the power of combining mosaic analysis with cell- or tissue-specific promoters is mentioned.
A genetic mosaic is an individual in which different cells have different genotypes. In Caenorhabditis elegans, genetic mosaics have been generated, identified, and analyzed for the purpose of defining where in the animal a gene of interest must be expressed to cause a particular phenotypic effect. Most C. elegans mosaics that have been studied were homozygous mutant for a recessive mutation in all cells and carried additionally, in some cells, one or more copies of the wild-type allele of the same gene on an extrachromosomal element. Before mosaic analysis is undertaken, a mutant phenotype corresponding to the absence of the wild-type gene from all cells is analyzed and described. Some mutant phenotypes are described in whole-animal terms, such as uncoordinated movement, longer-than-normal lifespan, abnormal body shape, or inviability. Such traits can be analyzed in mosaic animals. The overall phenotypes are recorded for mosaic animals in which it is known which cells carry the wild-type gene and which do not, with the goal of establishing which cells must carry the wild-type gene to prevent the appearance of the mutant phenotype. The responsible cell or group of cells is referred to as the anatomical focus of the action of the gene with respect to the phenotype under study.
In a second class of mutants, it is known at the outset that particular cells—specific nerve or muscle cells, for example—exhibit an abnormality, which is generally detected microscopically. Mosaic animals can then be used to ask whether or not the cellular focus of action with respect to this phenotype is in the very cell or cells that exhibit the abnormality; this is equivalent to asking whether or not the gene acts cell autonomously with respect to the cellular phenotype. When a gene acts cell autonomously, the phenotype of a cell is determined solely by the genotype of that cell and not by the genotypes of neighboring or other cells. A gene is said to act cell nonautonomously when a genotypically homozygous mutant cell exhibits a wild-type phenotype or when a cell carrying a wild-type allele exhibits a mutant phenotype. Cell non-autonomy is caused by intercellular interactions, and mosaic analysis can help elucidate the nature of the interaction. For example, a gene that produces a ligand that must be recognized by a receptor on a second cell could act cell nonautonomously with respect to a phenotypic effect in the receiving cell.
The generation, identification, and analysis of genetic mosaics in C. elegans have recently been reviewed (Herman, 1995; Yochem et al., 2000; Yochem and Herman, 2003). Also, a few of the technical details concerning mosaics are mentioned elsewhere in WormBook (see Nomarski images for learning the anatomy, with tips for mosaic analysis). Thus, these issues shall only be briefly mentioned here. Worm genetic mosaics have usually been generated from zygotes that were homozygous mutant for the gene under study and carried the wild-type allele of the same gene on an extrachromosomal element—either an extrachromosomal array or a free duplication of part of a chromosome—that was subject to spontaneous mitotic loss. If the extrachromosomal element is present in all cells, the animal exhibits a wild-type phenotype. But if the extrachromosomal element fails to be transmitted to a daughter cell during the development of the animal, all of the descendants of that cell, a clone, will lack the wild-type allele and be homozygous mutant. The extrachromosomal element should carry a second gene that behaves cell autonomously and can be scored to track which cells carry the extrachromosomal element and which do not. The well-described and nearly invariant cell lineage (Sulston et al., 1983) is a great aid in determining where in the lineage the extrachromosomal element was lost. This allows one to determine precisely, without scoring every cell, the genotypes of essentially every cell in the mosaic animal. Although the primary method has been described above, C. elegans mosaics have also been generated by other methods (Herman, 1995), and chimeric partial embryos have been formed in vitro from EMS and P2 blastomeres for genetic studies of Wnt signaling during early development (Thorpe et al., 1997; Schlesinger et al., 1999).
Over 75 genes have been analyzed in C. elegans genetic mosaics. Several examples of how mosaic analysis has contributed important insights into worm development were recently reviewed (Yochem et al., 2000; Yochem and Herman, 2003). Included were genes implicated in cell-to-cell signaling, with the functions of some genes assigned to the signaling cell and the functions of others assigned to the signal-receiving cell. Other examples in which anatomical foci were determined included genes affecting whole-animal phenotypes such as uncoordination, body size, and lifespan. A few additional examples are noted here.
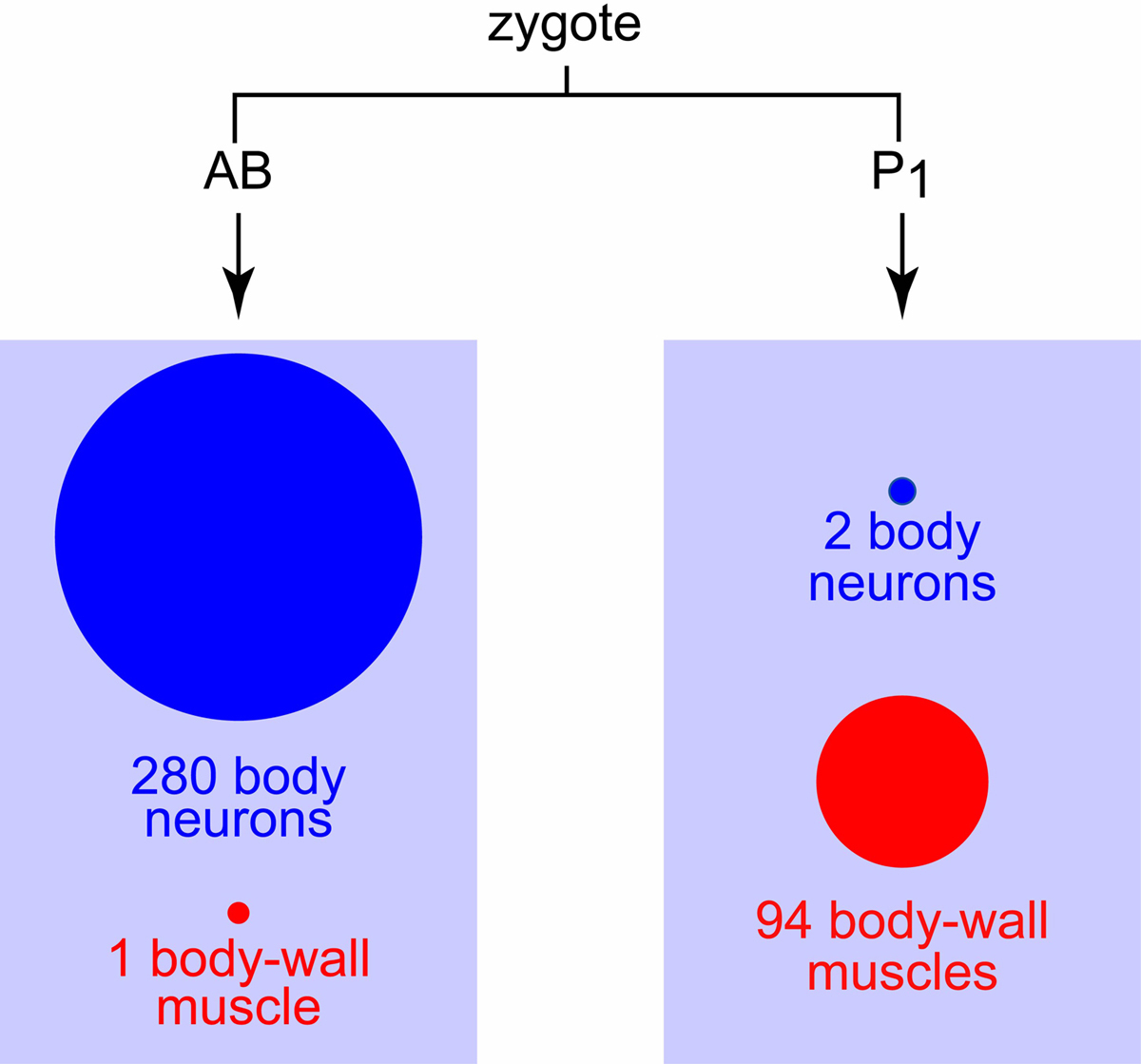
Mutations that confer uncoordinated movement often affect the body-wall muscles or the nervous system. The nature of the cell lineage makes it possible quickly to distinguish these candidate cell types: all of the body neurons, except for two, descend from AB, a daughter of the zygote; and all of the body-wall muscles, with the exception of one, descend from P1, the other daughter of the zygote (Sulston et al., 1983; Figure 1). A thorough analysis of mosaics is, or course, required for rigorous determination of the actual focus of activity of the gene, which could be in a cell type other than muscle or neuron. In a recent analysis, the anatomical focus of unc-122 has been traced to muscle (Loria et al., 2004). The unc-71 gene, which regulates both motor-axon guidance and sex-myoblast migration, is thought to act cell nonautonomously for both functions. The gene is required in both neurons and epidermis for proper motor-axon guidance (Huang et al., 2003).
|
Figure 1. AB-P1 mosaics with respect to body neurons and body-wall muscle. The separation of the precursors for most of the body neurons from those of most of the body-wall muscle occurs at the first division of the zygote (Sulston et al., 1983).
A problem for developmental studies is the use of signal-transduction molecules in several distinct processes. In some cases, later requirements for such molecules can be masked by a critical requirement for the molecules earlier in development. Mosaics can in some cases uncover the foci of activity of the gene in these various processes. A good example is mpk-1, which encodes an ERK2-type MAP-kinase known to affect vulval development (Lackner et al., 1994; Wu and Han, 1994). For proper vulval development, mpk-1 is required in the vulval precursor cells (Lackner et al., 1994). mpk-1 is also known to affect the exit of germ nuclei from the pachytene stage of meiosis, and in this case, the focus is the germ line itself (Church et al., 1995). The focus of a vital function for mpk-1 has now been shown to be hyp7, the large syncytium that composes most of the skin of the animal (Huang and Stern, 2004).
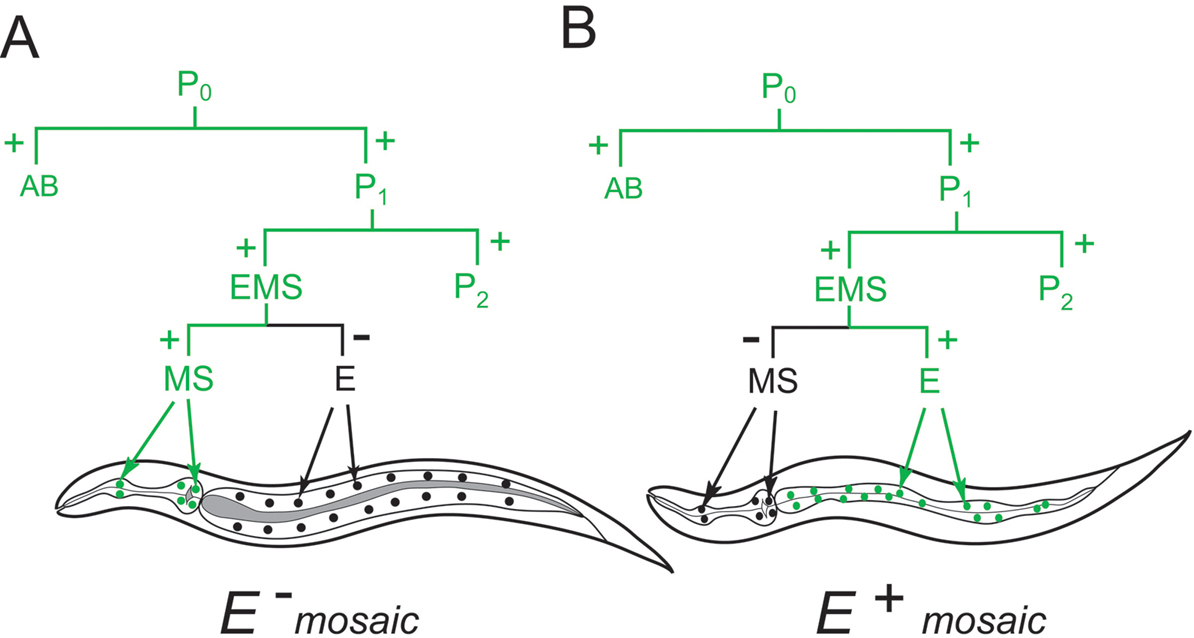
Illustrating the power of mosaics for non-developmental studies is bre-5, mutation of which confers resistance to a toxin from Bacillus thuringiensis (Griffitts et al., 2001). Animals expressing the dominant wild-type gene (conferring sensitivity to toxin) in the intestine were sensitive to the toxin, whereas 67 mosaics that were resistant to the toxin were always homozygous mutant in all of the cells of the intestine (Figure 2). The bacterial toxin appears to be harmful to C. elegans only if it can be ingested, which may require binding to a carbohydrate expressed by the intestinal cells. bre-5, which encodes a β-1,3-galactosyltransferase, may be required for synthesis of the carbohydrate (Griffitts et al., 2001).
|
Figure 2. Mosaicism and drug resistance or susceptibility resulting from loss of a bre-5+ extrachromosomal array during division of the blastomere EMS in an otherwise bre-5 mutant background. Inheritance of the array during development is inferred from expression of a green marker also present on the array. During development of the embryo in A, the array was inherited by MS but not by E, its sister. MS, the progenitor of several mesodermal cell types (a few in the pharynx are indicated), is not a progenitor of intestinal cells. They, instead, all derive from E, which only gives rise to intestinal cells (Sulston et al., 1983). Mosaics, such as the example in A, that have not inherited the array in the intestine (black nuclei), are resistant to the Bacillus thuringiensis toxin, indicating that bre-5 is required in the intestine for susceptibility (Griffitts et al., 2001). In contrast, the intestinal cells in mosaic B should express wild-type bre-5, because they have inherited the array (green nuclei). The presence of wild-type bre-5 in the gut makes the animal competent to ingest the toxin, resulting in intestinal pathology.
Potential problems with mosaic analysis have been mentioned in more detail in the recent reviews cited above and are only briefly mentioned here. Perhaps the foremost problem with mosaic analysis in C. elegans is that it requires detailed knowledge of the anatomy and cell lineage. Another problem is the potential for a cell that fails to inherit a wild-type gene carried by an extrachromosomal element nonetheless to receive some wild-type gene product made earlier in a progenitor cell that inherited the wild-type gene. Such perdurance of the gene product might weaken the expected mutant phenotype even when the cells that constitute the anatomical focus do not contain the wild-type gene.
A potential complication for mosaic analysis derives from the nature of the worm cell lineage. This is generally not a problem when investigating a cellular phenotype because mosaics can usually be identified in which the cell under investigation is genotypically different from cells that are candidates for interaction. The cell lineage can be limiting, however, when a focus of action of a gene is diffuse and distributed among cells of disparate lineage. The fully mutant phenotype may then only be apparent in mosaic animals when the duplication or array is lost by a progenitor of all or nearly all of the responsible cells, in which case it may be difficult to pinpoint the responsible cell types. A particularly difficult tissue is hyp7, a syncytium formed by the fusion of many mononucleate cells that descend from both AB and P1, the daughters of the very first embryonic cleavage (Sulston et al., 1983); therefore, no mosaic animal can contain a single, completely mutant hyp7 clone. Mosaic analyses have nonetheless been used to implicate hyp7 as the focus of action for several genes. Recent examples are unc-52 (Spike et al., 2002); mpk-1, which was mentioned above (Huang and Stern, 2004); genes of the TGFβ pathway (Wang et al., 2002); and tsp-15, which encodes a tetraspanin that affects epidermal morphology (Moribe et al., 2004).
The activity of a gene in a particular cell or tissue may be missed if the gene functions redundantly with another gene in that cell or tissue (see Genetic enhancers and Gene duplications and genetic redundancy in C. elegans). One may be suspicious if a GFP or other reporter construct indicates expression in a particular cell but genetic mosaics do not support an actual function. Of course, a reporter construct can give spurious results, if, for instance, cis-acting negative regulators are missing from the construct, and bona fide expression of a gene in a particular cell does not establish that the gene actually functions in that cell. If a redundant gene is identified, mosaic analysis can be performed in a background that is mutant for the redundant gene and vice versa. We are not aware that such analyses have yet been done in C. elegans, but in principle, they should be possible. An intriguing possibility is that two genes that redundantly affect a structure or function will have different foci with respect to that structure or function.
Mosaic analysis can also be difficult if the phenotype of a mutation is not completely penetrant. The question then arises whether a mosaic lacking the mutant phenotype is rescued in the appropriate cell type or is simply an escaper that has coincidental mosaicism. Analysis of a leaky mutation therefore requires caution and many mosaics.
In expression studies, some have no doubt encountered the situation in which a reporter construct rescued the mutant phenotype, but expression of the reporter gene could not be reliably detected. In some cases, antibodies against the reporter can provide increased sensitivity, but the concern remains that expression of a reporter construct gives spurious results, as mentioned above. Mosaic analysis, on the other hand, provides a functional test of the gene that does not depend on the ability to detect a reporter. Although there are potential limitations, as discussed above, mosaic analysis in many cases can provide an unbiased analysis of the activity of a gene, and the analysis can provide insight into the function of genes that are not easily amenable to immunocytochemical and other studies, because they are expressed at low levels in only a few cells.
A maternal contribution of a gene can be assessed when germ-line clones are generated following the loss of an extrachromosomal element in P1, P2, P3, or P4, the precursors of the germ cells (Sulston et al., 1983). A maternal contribution is evident when the progeny of a germ-line mosaic have a more severe phenotype than progeny that segregate from a mother who is heterozygous for the gene (for an example, see Herman, 1995). A full-fledged mosaic analysis is not necessary for an analysis of germ-line clones. They can be seen in the course of maintaining lines rescued by an extrachromosomal element. If many individuals are put one to a plate, a few of the plates may have received a germ-line mosaic. For a gene with a completely penetrant phenotype, all of the progeny will have a mutant phenotype. A maternal contribution can also be assessed with RNA interference (RNAi), although the potential for cross-interference and for incomplete depletion of gene product should be kept in mind.
Another method for asking about the anatomical focus of the activity of a gene is to introduce transgenic copies of the wild-type gene controlled by cell- or tissue-specific promoters into otherwise homozygous mutant animals. Rescue of a mutant defect by a particular transgene indicates that expression in the indicated cell type can provide the needed function. There are, however, some concerns when using this approach. One is the fidelity of expression of such transgenes, especially from arrays that contain multiple copies of the engineered gene and other heterologous DNA. Also, a bit of gene cloning is involved, because a new construct must be made for each promoter under study. The formal possibility also remains that rescue by expression (or overexpression) from a transgene does not prove that the wild-type gene normally acts in the same cell type or tissue. Nevertheless, this approach, especially in combination with mosaic analysis, is very powerful, as recent examples demonstrate (Zhen and Jin, 1999; Inoue and Thomas, 2000; Zhen et al., 2000; Wang et al., 2002; Huang et al., 2003; Huang and Stern, 2004; Loria et al., 2004). Such combined analyses should continue to provide some of the most convincing insight into the cell specificity of gene function in C. elegans.
Church, D.L., Guan, K.L., and Lambie, E.J. (1995). Three genes of the MAP kinase cascade, mek-2, mpk-1/sur-1 and let-60 ras, are required for meiotic cell cycle progression in Caenorhabditis elegans. Development 121, 2525–2535. Abstract
Griffitts, J.S., Whitacre, J.L., Stevens, D.E., and Aroian, R.V. (2001). Bt toxin resistance from loss of a putative carbohydrate-modifying enzyme. Science 293, 860–864. Abstract Article
Herman, R.K. (1995). Mosaic analysis. Methods Cell Biol. 48, 123–146. Abstract
Huang, P., and Stern, M.J. (2004). FGF signaling functions in the hypodermis to regulate fluid balance in C. elegans. Development 131, 2595–2604. Abstract Article
Huang, X., Huang, P., Robinson, M.K., Stern, M.J., and Jin, Y. (2003). UNC-71, a disintegrin and metalloprotease (ADAM) protein, regulates motor axon guidance and sex myoblast migration in C. elegans. Development 130, 3147–3161. Abstract
Inoue, T., and Thomas, J.H. (2000). Targets of TGF-β signaling in Caenorhabditis elegans dauer formation. Dev. Biol. 217, 192–204. Abstract Article
Lackner, M.R., Kornfeld, K., Miller, L.M., Horvitz, H.R., and Kim, S.K. (1994). A MAP kinase homolog, mpk-1, is involved in ras-mediated induction of vulval cell fates in Caenorhabditis elegans. Genes Dev. 8, 160–173. Abstract
Loria, P.M., Hodgkin, J., and Hobert, O. (2004). A conserved postsynaptic transmembrane protein affecting neuromuscular signaling in Caenorhabditis elegans. J. Neurosci. 24, 2191–2201. Abstract Article
Moribe, H., Yochem, J., Yamada, H., Tabuse, Y., Fujimoto, T., and Mekada, E. (2004). Tetraspanin protein (TSP-15) is required for epidermal integrity in Caenorhabditis elegans. J. Cell Sci. Pt. Abstract Article
Schlesinger, A., Shelton, C.A., Maloof, J.N., Meneghini, M., and Bowerman, B. (1999). Wnt pathway components orient a mitotic spindle in the early Caenorhabditis elegans embryo without requiring gene transcription in the responding cell. Genes Dev. 13, 2028–2038. Abstract
Spike, C.A., Davies, A.G., Shaw, J.E., and Herman, R.K. (2002). MEC-8 regulates alternative splicing of unc-52 transcripts in C. elegans hypodermal cells. Development 129, 4999–5008. Abstract
Sulston, J.E., Schierenberg, E., White, J.G., and Thomson, J.N. (1983). The embryonic cell lineage of the nematode Caenorhabditis elegans. Dev. Biol. 100, 64–119. Abstract Article
Thorpe, C.J., Schlesinger, A., Carter, J.C., and Bowerman, B. (1997). Wnt signaling polarizes an early C. elegans blastomere to distinguish endoderm from mesoderm. Cell 90, 695–705. Abstract Article
Wang, J., Tokarz, R., and Savage-Dunn, C. (2002). The expression of TGFβ signal transducers in the hypodermis regulates body size in C. elegans. Development 129, 4989–4998. Abstract
Wu, Y., and Han, M. (1994). Suppression of activated Let-60 ras protein defines a role of Caenorhabditis elegans Sur-1 MAP kinase in vulval differentiation. Genes Dev. 8, 147–159. Abstract
Yochem, J., and Herman, R.K. (2003). Investigating C. elegans development through mosaic analysis. Development 130, 4761–4768. Abstract Article
Yochem, J., Sundaram, M., and Bucher, E.A. (2000). Mosaic analysis in Caenorhabditis elegans. Methods Mol. Biol. 135, 447–462. Abstract
*Edited by Jonathan Hodgkin and Philip Anderson. Last revised March 14, 2005. Published December 27, 2005. This chapter should be cited as: Yochem, J. and Herman, R. K. Genetic mosaics (December 27, 2005), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.58.1, http://www.wormbook.org.
Copyright: © 2005 John Yochem and Robert K. Herman. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
§To whom correspondence should be addressed. E-mail: yoche001@umn.edu
 All journal content, except where otherwise noted, is licensed under a Creative Commons Attribution License.
All journal content, except where otherwise noted, is licensed under a Creative Commons Attribution License.