Embryo series courtesy of Einhard Schierenberg
Embryo series courtesy of Einhard SchierenbergTable of Contents
Abstract
In the last decade, nematodes other than C. elegans have been studied intensively in evolutionary developmental biology. A few species have been developed as satellite systems for more detailed genetic and molecular studies. One such satellite species is the diplogastrid nematode Pristionchus pacificus. Here, I provide an overview about the biology, phylogeny, ecology, genetics and genomics of P. pacificus.
P. pacificus was described as a novel species in 1996 and the first isolate PS312 from Pasadena (California) was established as a laboratory strain (Sommer et al., 1996; Sommer and Sternberg, 1996). By now, 15 strains from three different continents (North America, Europe and Asia) are in culture, some of which differ molecularly and developmentally from one another (Schlak et al., 1997; Srinivasan et al., 2001).
In the last ten years, P. pacificus has been established as a satellite organism in evolutionary developmental biology (Hong and Sommer, 2006). P. pacificus is a self-fertilizing hermaphrodite, has a 4-day life cycle at 20°C and can be cultured on OP50. P. pacificus is amenable to various cellular, genetic and molecular techniques successfully used in C. elegans. In addition to forward genetics, morpholino knockdown and deletion library experiments provide reverse genetic tools (for genetic and genomic methods see P. pacificus genetics and P. pacificus genomics chapters in WormMethods; Zheng et al., 2005).
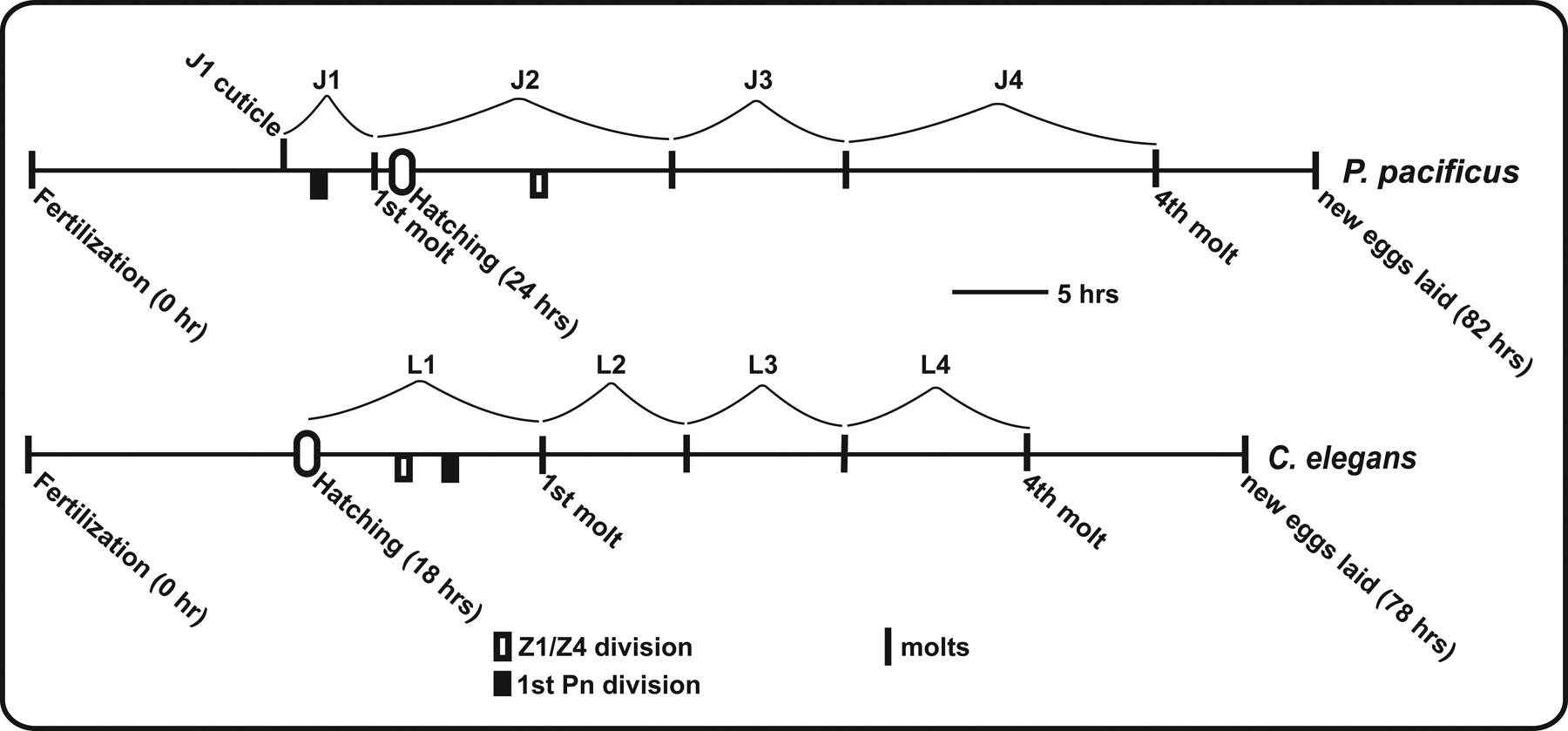
Under laboratory conditions, the life cycle of P. pacificus is nearly as fast as the one of C. elegans, but differs in one important feature: P. pacificus, like all species of the family Diplogastridae, has an embryonic molt. J1 larvae molt to J2 before they hatch from the egg (Figure 1; Fürst v. Lieven, 2005). Thus, the J1 stage is not free-living and non-feeding. It has been suggested that the non-feeding J1 stage allows for a complex stoma morphology compared to the basal rhabditid buccal structure. Indeed, some taxa of the Diplogastridae (including P. pacificus) show an interesting dimorphism of stoma structures (Fürst von Lieven & Sudhaus, 2000). The two alternative mouth forms are called eurystomatous and stenostomatous morph, respectively (Figure 2; Fürst von Lieven & Sudhaus, 2000). The stenostomatous buccal cavity is narrow, whereas eurystomatous worms have a broad buccal cavity. Among the different tooth-like structures present in the buccal cavity, eurystomatous worms have a claw-like dorsal tooth and a specialized tooth in the right subventral sector acting as antagonist, while stenostomatous worms lack the tooth of the right subventral side, and the dorsal tooth is not claw-like as in eurystomatous worms (Figrue 2). These differences in buccal morphology are though to be related to different feeding habits (Fürst von Lieven & Sudhaus, 2000). Indeed, P. pacificus can feed on various bacteria, fungi and even other nematodes.
|
Figure 1: Life cycle of P. pacificus in comparison to C. elegans. P. pacificus propagates through four juvenile stages, called J1 to J4. In contrast to C. elegans, the J1 to J2 molt is embryonic and only the J2 stage hatches from the egg. Dauer formation occurs as alternative J3 stage, as in C. elegans. The direct life cycle takes approximately 4 days at 20°C. Reprinted with permission from Hong and Sommer (2006), Bioessays © copyright 2006 Wiley Periodicals, Inc.
|
Figure 2: Dimorphism of the buccal cavity. Dorsal is left. (a) Stenostomatous mouth form, which is slightly deeper than wide (compare the distance between the parallel dashed lines and the width of the buccal cavity) and a flint-like dorsal tooth (arrowhead). (b-c) Eurystomatous mouth form with a broader buccal cavity, showing the same individual at different focal planes. Notice a claw-like dorsal tooth (b, arrow head), and an adjacent row of denticles (c, arrow). Scale bar represents 5 μm in all panels. Reprinted with permission from Hong and Sommer (2006), Bioessays © copyright 2006 Wiley Periodicals, Inc.
The first developmental process to be studied in great detail in P. pacificus was vulva formation (see Evolution of development in nematodes related to C. elegans). When compared to C. elegans, vulva development in P. pacificus involves a set of evolutionary modifications, including i) programmed cell death of non-vulval epidermal cells P(1-4,9-11).p, ii) vulva induction by multiple cells of the somatic gonad and iii) novel cell-cell interactions during vulva formation (see Evolution of development in nematodes related to C. elegans). P. pacificus vulva defective mutants have been isolated in large-scale mutagenesis screens and the phenotypes of mutations have helped elucidating the molecular mechanisms of evolutionary change. For example, mutations in Ppa-lin-17/Frizzled result in gonad-independent vulva differentiation and a multivulva phenotype indicating a role of Wnt signaling in a negative signaling process (Zheng et al., 2005). Besides vulva formation, gonad development (Rudel et al., 2005), sex determination (Pires-daSilva & Sommer, 2004), mesoderm development (Photos et al., 2006), dauer formation, olfaction and other aspects of neurobiology are studied intensively in P. pacificus.
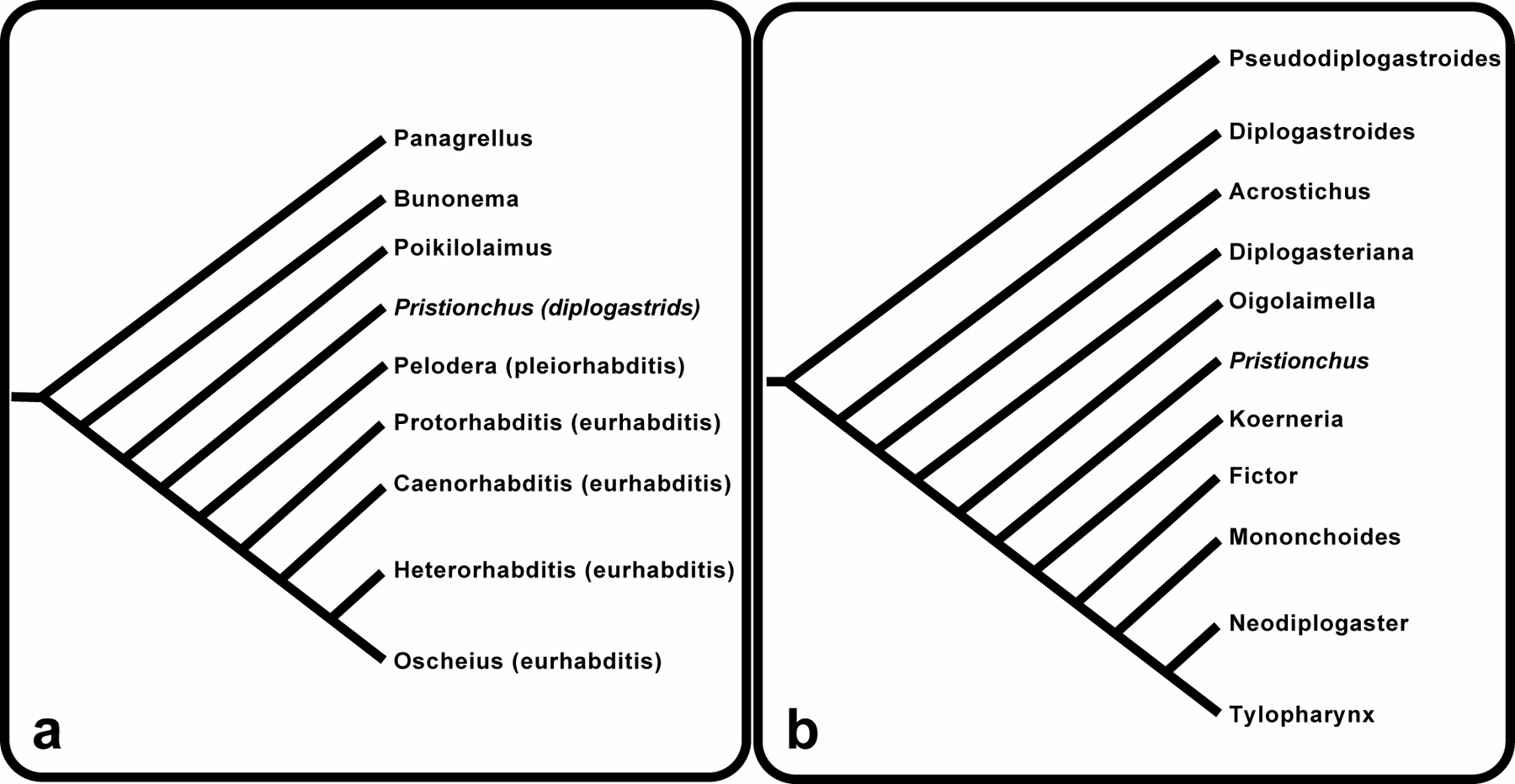
P. pacificus belongs to the family Diplogastridae, which includes about 300 described free-living species in 28 genera (Fürst v. Lieven & Sudhaus, 2000; Sudhaus & Fürst v. Lieven, 2003). The exact phylogenetic relationship to the Rhabditidae is still debated and several studies suggest that the Diplogastridae are part of the Rhabditidae (Figure 3a; see also Figure 1 in The phylogenetic relationships of Caenorhabditis and other rhabditids). Within the Diplogastridae, the genus Pristionchus represents a more derived taxon (Figure 3b). However, more molecular studies are necessary to provide a clear picture of the phylogenetic relationship of diplogastrid genera.
|
Figure 3: Nematode phylogeny. (a) Phylogeny of free-living nematodes based on the analysis of nucleotide sequence data. (b) Diplogastridae phylogeny based on morphological apomorphies of each genus (adapted from Sudhaus and Lieven 2003). Reprinted with permission from Hong and Sommer (2006), Bioessays © copyright 2006 Wiley Periodicals, Inc.
Considering that the whole genome sequence comparison between C. elegans and C. briggsae suggests a separation time of 80–120 million years for those two species, the separation time of P. pacificus and C. elegans is expected to be much older. However, as no fossil record exists for nematodes, no accurate numbers can be given. We are currently working on accurate estimates within the genus Pristionchus by using detailed molecular phylogenetics (W. Mayer and R. J. Sommer, ongoing studies).
Little is known about the ecology of many so-called “free-living” nematode species. Only very recently several studies found that C. elegans occurs predominantly in compost heaps (see Natural variation and population genetics of Caenorhabditis elegans and Ecology of Caenorhabditis species). In Pristionchus, recent studies revealed that members of this genus live in close association with scarab beetles and the Colorado potato beetle (Herrmann et al., 2006). Sampling of beetles in Western Europe in the years 2004 and 2005 resulted in more than 350 isolates that fell into six species. Two hermaphroditic species, P. entomophagus and P. maupasi accounted for 60% of the isolates and occurred on dung beetles and cockchafers, respectively. However, the satellite organism P. pacificus was neither observed on scarab beetles nor on the Colorado potato beetle in Western Europe.
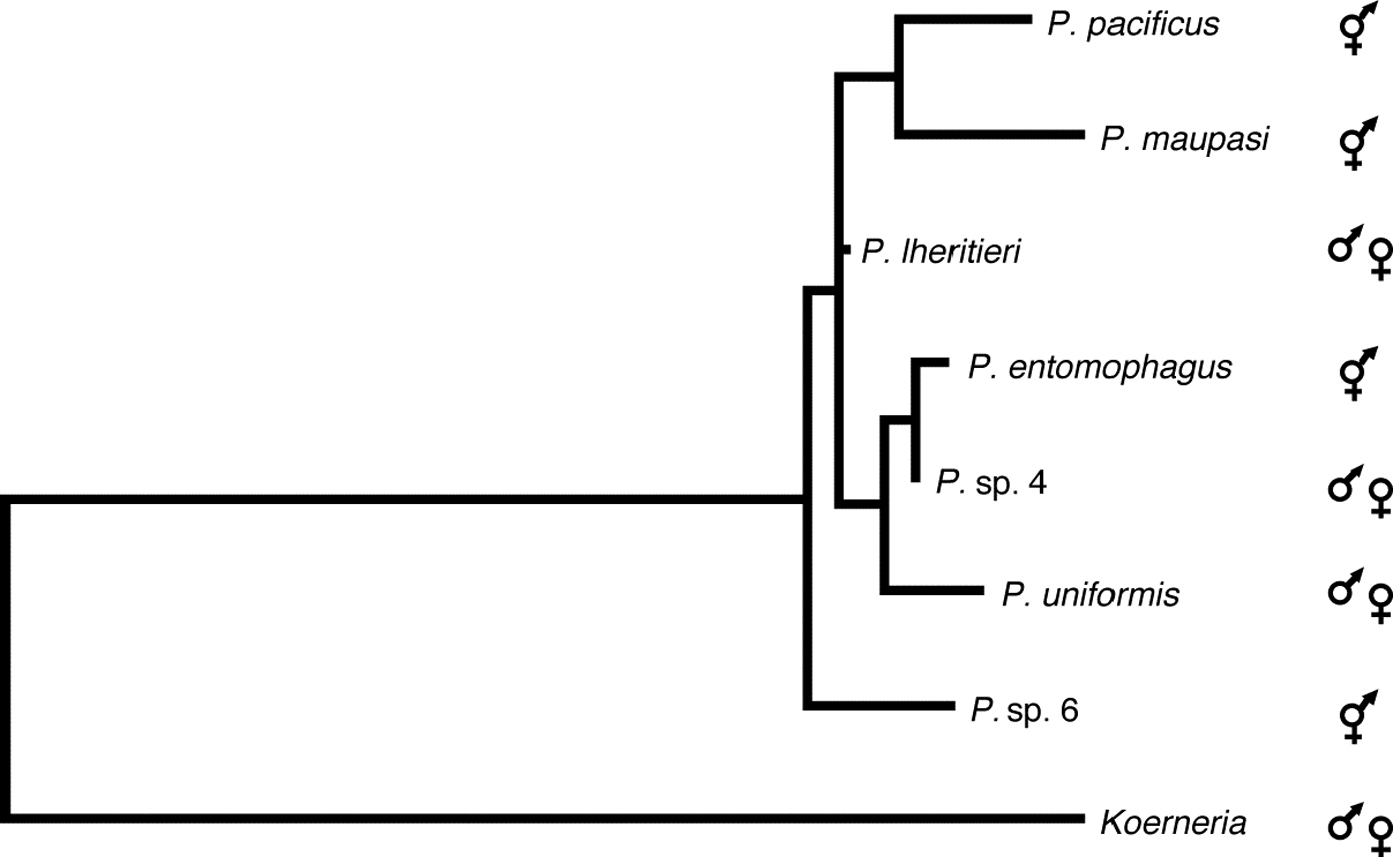
Similar studies of Pristionchus species associated with scarab beetles and the Colorado potato beetle in the Eastern United States revealed striking differences in the species composition and mode of reproduction of Pristionchus species between Europe and North America (Herrmann, Mayer, & Sommer, unpublished). While 60% of the wild isolates from Europe belong to two hermaphroditic species, more than 95% of the American isolates belong to several gonochoristic species. These results establish Pristionchus as a nematode model system for speciation, biogeography and biodiversity. In total, 15 Pristionchus species are available in culture and are used for studying microevolution of developmental processes, olfaction and genomics. The phylogenetic relationship of seven of these 15 species is represented in Figure 4.
|
Figure 4: Pristionchus phylogeny. Phylogenic maximum likelihood tree based on SSU sequences of the genus Pristionchus. The sequence of Koerneria sp. was included as the closest related genus to Pristionchus. The phylogenetic relationship was inferred by the heuristic search algorithm of the PAUP*4.0b10 program using the default settings under the maximum likelihood criterion. The tree was rooted at midpoint. Hermaphroditic and gonochoristic species are indicated. Numbers at nodes indicate bootstrap values after 1000 replications. Reprinted from Herrmann et al., (2006), with permission from Elsevier.
P. pacificus is a self-fertilizing hermaphrodite with the frequent occurrence of males. The sex determination system is of the XX/XO type, similar to C. elegans (Pires-daSilva & Sommer, 2004). In general, mutagenesis experiments are carried out in the laboratory strain P. pacificus PS312 from California and can be performed using various mutagens, such as EMS or TMP/UV. However, RNAi and transformation methods are still being developed. The P. pacificus genetics chapter in WormMethods by Pires-daSilva provides detailed protocols for various forward - and reverse genetic approaches in P. pacificus. For formal genetics, a set of morphological mutants is available (Kenning et al., 2004). Most of these mutants have Dumpy-like phenotypes, but mutants with Unc-like phenotypes are available as well.
In general, the C. elegans nomenclature was adopted for P. pacificus by adding the prefix “Ppa” to indicate the species. To prevent confusion between individual gene names in P. pacificus and C. elegans the following rules are used to distinguish phenotypic classes of mutants and orthologous genes: Genetically defined mutants in P. pacificus are described with a prefix and a novel abbreviation, i.e. Ppa-pdl-n, with “pdl” standing for “Pristionchus-dumpy-like”. This type of nomenclature system is chronological. Generally, mutants are re-named after the molecular lesion of a gene has been identified and was shown to be orthologous to a known C. elegans gene. On the other hand, molecularly defined orthologs of C. elegans genes are described as Ppa-ortholog, i.e. Ppa-lin-39 and Ppa-mab-5 for the Hox genes (Eizinger and Sommer, 1997; Jungblut and Sommer, 1998).
A genomic initiative in P. pacificus was launched in 2001. An integrated genome map of P. pacificus contains a genetic linkage map of more than 500 molecular markers and a physical map of nearly 10.000 fingerprinted BAC clones (Srinivasan et al., 2002, Srinivasan et al., 2003). A whole-genome sequencing project is ongoing (http://www.nhgri.nih.gov/12511858) and should result in a high-coverage draft within 2006.
The comparison of the genetic linkage maps of P. pacificus and C. elegans revealed that, with one exception, all chromosomes can easily be homologized (Table 1). The P. pacificus chromosomes II, III, IV and X are homologous to the corresponding chromosomes of C. elegans. P. pacificus chromosome V corresponds to C. elegans chromosome I and vice versa. However, one arm of P. pacificus chromosome I contains genes that are orthologous to genes of the C. elegans X chromosome indicating one major chromosomal translocation after the split of Pristionchus and Caenorhabditis (Table 1; Srinivasan et al., 2002).
Table 1: Comparisons of P. pacificus and C. elegans chromosomes and their total genetic distances. Reprinted with permission from Hong and Sommer (2006), Bioessays © copyright 2006 Wiley Periodicals, Inc.
| Number of Ppa SNP markers | P. pacificus chromosome | C. elegans chromosome | Ppa_chr genetic distances (cM) | Cel_chr genetic distances (cM) |
|---|---|---|---|---|
| 110 | I | V, X | 215 | 50 |
| 76 | II | II | 168 | 50 |
| 78 | III | III | 117 | 50 |
| 78 | IV | IV | 179 | 50 |
| 72 | V | I | 162 | 50 |
| 65 | X | X | 185 | 50 |
Although these data prove the existence of macro-synteny, other genetic and physical properties differ between the P. pacificus and C. elegans genomes. Genetically, the strongest difference between the two organisms is the absence of interference in P. pacificus. Interference describes the genetic phenomenon that the first chiasmata during meiosis at one particular chromosome inhibits the occurrence of another chiasmata along the same chromosome. C. elegans shows nearly complete interference, resulting in one cross-over per chromosome per meiosis. By definition, C. elegans chromosomes are nearly 50 cM in length. In P. pacificus double cross-overs are frequent and most chromosomes are genetically much larger than 50 cM. In total, the P. pacificus genome has a length of 1026 cM with the caveat that the distal-most markers have not been physically linked to the telomeres yet (Table 1).
In addition to the larger genetic size of the P. pacificus genome, the physical size also differs from the one in C. elegans. Fluoremetric measurements indicated a size of 161 Mb (Prof. S. Johnston, pers. Communication), a value that awaits further confirmation by the ongoing sequencing project.
When genomes of related organisms are compared, the level of conservation of gene order can be expressed as synteny. The term microsynteny indicates that the order of individual genes in considered genomes is exactly conserved. BAC clone sequence analysis, as well as the ongoing whole genome sequencing initiative, revealed that the degree of microsynteny between the genomes of P. pacificus and C. elegans is limited (Lee et al., 2003). The complete sequence of a BAC clone 7E22 of P. pacificus provided a continuous sequence of 126 kb. It contains a region flanking the caudal-homolog pal-1 of P. pacificus and contains 20 predicted open reading frames (ORFs), 11 of which have putative orthologs in C. elegans. 10 of these 11 orthologs are located on C. elegans chromosome III. However, most of these genes are distributed over more than a 12 Mb interval of the C. elegans genome and only three pairs of genes show microsynteny (Lee et al., 2003).
In addition, the comparison between P. pacificus and C. elegans reveals differences in the genetic repertoire of both species. For example, P. pacificus contains a dnmt-2-like DNA methyltransferase gene, whereas dnmt-2-like genes are completely absent from C. elegans and C. briggsae (Gutierrez and Sommer, 2004).
In C. elegans, up to 15% of the genes are organized in operons and polycistronic precursor RNAs are processed by trans-splicing at the 5′ ends of genes through the addition of a specific trans-spliced leader (see Trans-splicing and operons). Among the 10 different spliced leaders known from C. elegans, the SL1 leader is most abundant. The SL1 leader is spliced to the 5′ ends of monocistronic genes and to upstream genes in operons. Trans-splicing is common among nematodes and was observed in the genera Panagrellus, Ascaris, Haemonchus, Anisakis and Brugia (Bektesh et al., 1988). With regard to operons, the best available information is on Brugia malayi (Whitton et al., 2004).
In P. pacificus, SL1 trans-splicing occurs commonly and the SL1 leader sequence is identical to the SL1 leader of C. elegans (Jungblut and Sommer, 1998). In addition, P. pacificus contains operons, downstream genes of which are trans-spliced to SL2 (Lee and Sommer, 2003). Surprisingly, the operons analyzed so far in P. pacificus are not conserved in C. elegans.
The genetic linkage map with more than 500 molecular markers and the physical map provide useful tools for the mapping and cloning of P. pacificus mutants. For mapping, the laboratory strain P. pacificus PS312 from California is crossed with the mapping strain P. pacificus PS1843 from Washington. These two strains differ in approximately 3% of their sequence and provide large amounts of SNPs. The ongoing sequencing project provides a 1 X coverage of the P. pacificus Washington sequence, which allows the detection of potential polymorphisms by BLAST search.
Mapping strategies are different from C. elegans protocols given that the available markers are mostly SSCP markers. The P. pacificus genomics chapter in WormMethods by Srinivasan and Dieterich provides detailed protocols for the mapping procedures and sequence searches. All of this information is available at the P. pacificus database (http://www.pristionchus.org/cgi-bin/index.pl).
I thank Metta Riebesell for critically reading this manuscript and the Max-Planck Society for support of our research on Pristionchus pacificus.
Bektesh, S., Van Dore, K., and Hirsh, D. (1988). Presence of the Caenorhabditis elegans spliced leader on different mRNAs and in different genera of nematodes. Genes Dev. 2, 1277–1283. Abstract
Eizinger, A., and Sommer, R.J. (1997). The homeotic gene lin-39 and the evolution of nematode epidermal cell fates. Science 278, 452–455. Abstract Article
Fürst von Lieven, A., and Sudhaus, W. (2000). Comparative and functional morphology of the buccal cavitiy of Diplogastrina (Nematoda) and a first outline of the phylogeny of this taxon. J. Zoolog. Syst. Evol. Res. 38, 37–63. Article
Fürst von Lieven, A. (2005). The embryonic moult in diplogastrids (Nematoda) - homology of developmental stages and heterochrony as a prerequisite for morphological diversity. Zool. Anz. 244, 79–91.
Gutierrez, A., and Sommer, R.J. (2004). Evolution of dnmt-2 and mbd-2-like genes in the free-living nematodes Pristionchus pacificus, Caenorhabditis elegans and Caenorhabditis briggsae. Nucleic Acids Res. 32, 6388–6396. Abstract Article
Herrmann, M., Mayer, W., and Sommer, R.J. (2006). Nematodes of the genus Pristionchus are closely associated with scarab beetles and the Colorado potato beetle in western Europe. Zoology 109, 96–108. Abstract Article
Hong, R.L., and Sommer, R.J. (2006). Pristionchus pacificus: a well-rounded nematode. Bioessays 28, 651–659. Abstract Article
Jungblut, B., and Sommer, R.J. (1998). The Pristionchus pacificus mab-5 gene is involved in the regulation of ventral epidermal cell fates. Curr. Biol. 8, 775–778. Abstract Article
Kenning, C., Kipping, I., and Sommer, R.J. (2004). Mutations with altered gross-morphology in the nematode Pristionchus pacificus. Genesis 40, 176–183. Abstract Article
Lee, K.-Z., Eizinger, A., Nandakumar, R., Schuster, S.C., and Sommer, R.J. (2003). Limited microsynteny between the genomes of Pristionchus pacificus and Caenorhabditis elegans. Nucleic Acids Res. 10, 2553–2560. Abstract Article
Lee, K.-Z., and Sommer, R.J. (2003). Operon structure and trans-splicing in the nematode Pristionchus pacificus. Mol. Biol. Evol. 20, 2097–2103. Abstract Article
Photos, A., Gutierrez, A., and Sommer, R.J. (2006). sem-4/spalt and egl-17/FGF have a conserved role during sex myoblast specification and migration in P. pacificus and C. elegans. Dev. Biol. 293, 142–153. Abstract Article
Pires-daSilva, A., and Sommer, R.J. (2004). Conservation of the global sex determination gene tra-1 in distantly related nematodes. Genes Dev. 18, 1198–1208. Abstract Article
Rudel, D., Riebesell, M., and Sommer, R. J. (2005). Gonadogenesis in Pristionchus pacificus and organ evolution: development, adult morphology and cell/cell interactions in the hermaphrodite gonad. Dev. Biol. 277, 200–221. Abstract Article
Schlak, I., Eizinger, A., and Sommer, R.J. (1997). High rate of restriction fragment length polymorphisms between two populations of the nematode Pristionchus pacificus (Diplogastridae). J. Zoolog. Syst. Evol. Res. 35, 137–142.
Srinivasan, J., Pires-daSilva, A., Gutierrez, A., Zheng, M., Jungblut, B., Witte, H., Schlak, I. and Sommer, R.J. (2001). Microevolutionary analysis of the nematode genus Pristionchus suggests a recent evolution of redundant developmental mechanisms during vulva formation. Evol. Dev. 3, 229–240. Abstract Article
Srinivasan, J., Sinz, W., Lanz, C., Brand, A., Nandakumar, R., Raddatz, G., Witte, H., Keller, H., Kipping, I., Pires-daSilva, A., Jesse, T., Millare, J., de Both, M., Schuster, S.C., and Sommer, R.J. (2002). A BAC-based genetic linkage map of the nematode Pristionchus pacificus. Genetics 162, 129–134. Abstract
Srinivasan, J., Sinz, W., Jesse, T., Wiggers-Perebolte, L., Jansen, K., Buntjer, J., van der Meulen, M., and Sommer, R.J. (2003). An integrated physical and genetic map of the nematode Pristionchus pacificus. MGG 269, 715–722. Abstract
Sommer, R.J., Carta, L.K., Kim, S.-Y., and Sternberg, P.W. (1996). Morphological, genetic and molecular description of Pristionchus pacificus sp. n. (Nematoda, Diplogastridae). Fundam. Appl. Nematol. 19, 511–521.
Sommer, R.J., and Sternberg, P.W. (1996). Apoptosis limits the size of the vulval equivalence group in Pristionchus pacificus: a genetic analysis. Curr. Biol. 6, 52–59. Abstract Article
Sudhaus, W., and Fürst von Lieven, A. (2003). A phylogenetic classification and catalogue of the Diplogastridae (Secernentea, Nematoda). J. Nematode Morphol. System. 6, 43–90.
*Edited by Jonathan Hodgkin. Last revised May 25, 2006. Published August 14, 2006. This chapter should be cited as: Sommer, R.J. Pristionchus pacificus (August 14, 2006), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.102.1, http://www.wormbook.org.
Copyright: © 2006 Ralf J. Sommer. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
§To whom correspondence should be addressed. Phone: ++ 49 7071 601 371, Fax: ++ 49 7071 601 498, E-mail: ralf.sommer@tuebingen.mpg.de
 All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.
All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.