Embryo series courtesy of Einhard Schierenberg
Embryo series courtesy of Einhard SchierenbergTable of Contents
Abstract
The intestine is one of the major organs in C. elegans and is largely responsible for food digestion and assimilation as well as the synthesis and storage of macromolecules. In addition, the intestine is emerging as a powerful experimental system in which to study such universal biological phenomena as vesicular trafficking, biochemical clocks, stress responses and aging. The present chapter describes some of these many and varied properties of the C. elegans intestine: the embryonic cell lineage, intestine morphogenesis, structure and physiology of the intestinal cell and, finally, the transcription factor network controlling intestine development and function.
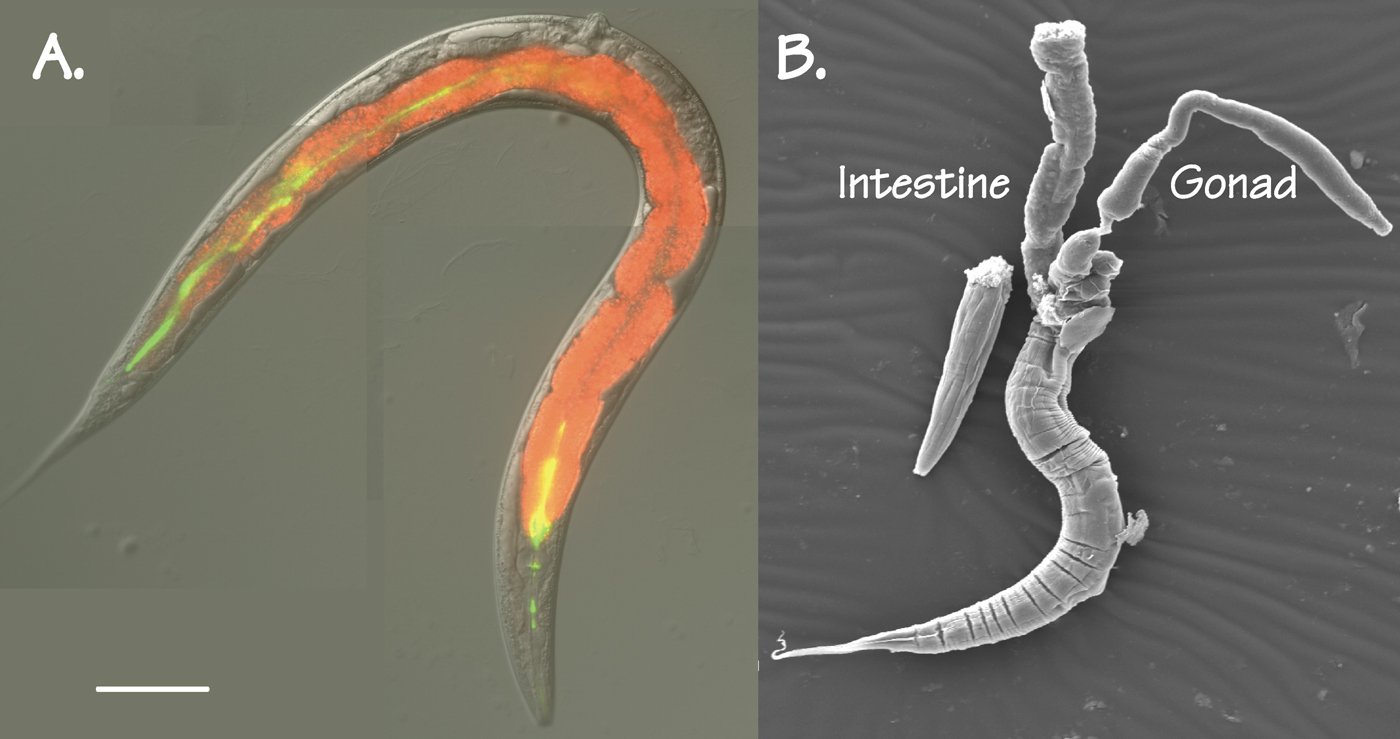
The intestine is one of the major organs of C. elegans, comprising roughly one third of the total somatic mass. Two views of the intestine are shown in Figure 1. Figure 1A is a differential interference contrast image of an adult C. elegans hermaphrodite (actually, a gonadless-mutant) on which have been superimposed two fluorescent images: Nile red to reveal fat droplets in the intestine cytoplasm and green fluorescent latex beads to reveal the intestine lumen. Figure 1B shows a scanning electron micrograph of an adult hermaphrodite, decapitated to allow the intestine (and a gonad) to spill from the carcass. Many additional views of the C. elegans intestine, especially transmission electron micrographs, can be inspected at Wormatlas.
|
Figure 1. Two views of the adult C. elegans intestine. A. An adult glp-4(bn2) C. elegans hermaphrodite, raised at the restrictive temperature to prevent formation of a gonad. Differential interference contrast image; red reveals fat droplets in the intestine (Nile Red Fluorescence); green reveals the presence of (indigestible) fluorescent polystyrene beads in the intestinal lumen. Scale bar = 0.1 mm.B. Scanning electron micrograph of a decapitated adult C. elegans hermaphrodite, showing intestine and gonad. (Photograph courtesy of Mr. Wei Dong, University of Calgary).
At the peak of egg laying, an adult hermaphrodite converts her body mass to embryos once per day (Hirsh et al., 1976), corresponding to several million bacteria consumed, and it is likely that the majority of this mass passes through the intestine (ignoring the possibility of gas or water exchange across the cuticle). Considering that the average residence time for a bacterium within the intestine is less than two minutes (S. Ghafouri and JDM, in press; see also Avery and Shtonda (2003)), the digestive and assimilative power of the worm intestine must be enormous. The present chapter aims to describe the varied aspects of intestinal biology in C. elegans, beginning with the intestinal cell lineage, development and morphogenesis, and then proceeding to the cell biology of the polarized enterocyte, to the wide range of biochemical reactions that are centred in the intestine and finally, to the transcription factor network that regulates all of these genes. Wherever possible, reference will be made to other chapters in the Wormbook that are more tightly focused on particular biological processes. The term “intestine” will be used in preference to “gut”; in other metazoons, “gut” usually refers to the entire digestive tract (pharynx, intestine and rectum, for C. elegans).
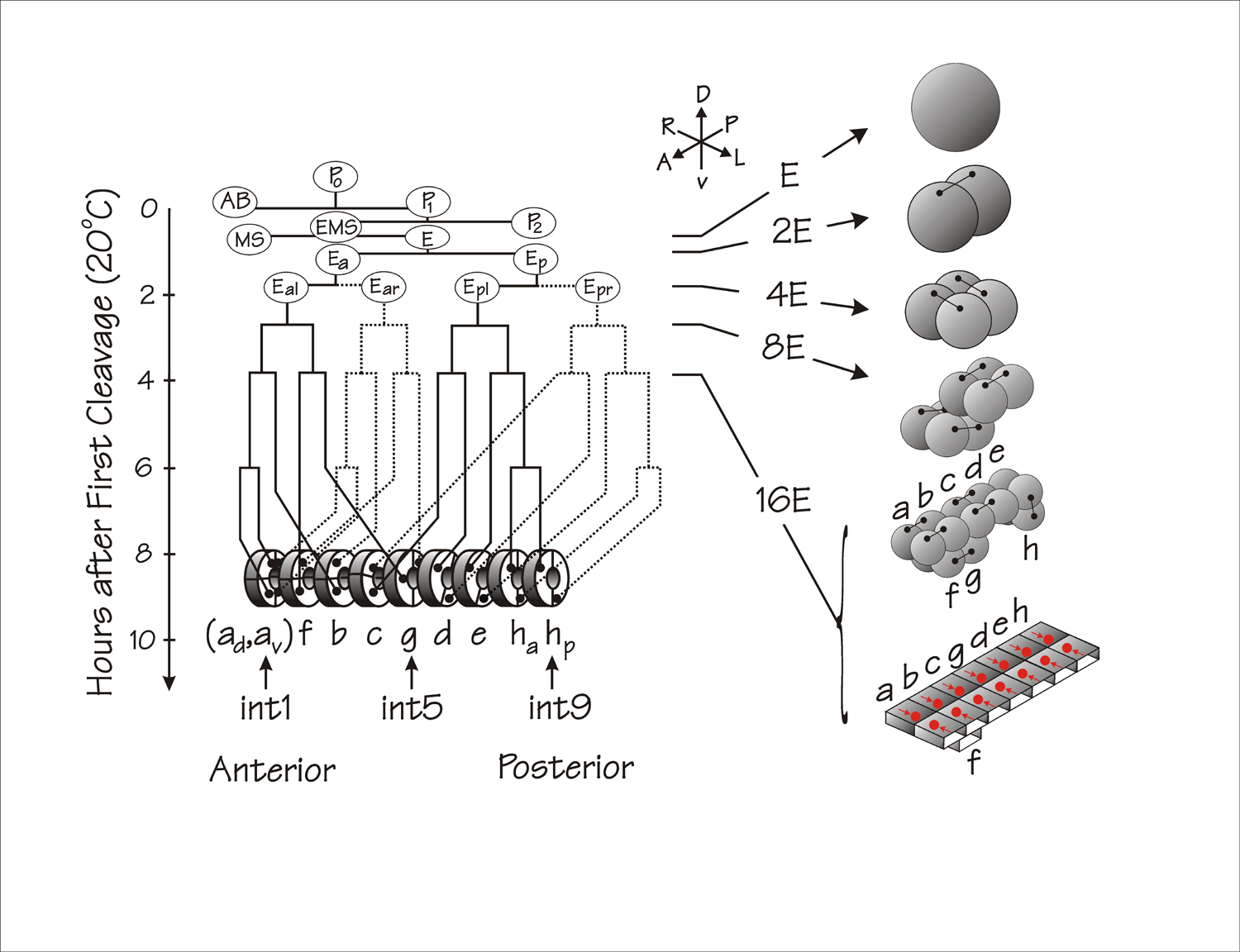
The entire intestine is produced as a clone of cells descending from a single cell, the E blastomere, of the eight-cell embryo (Deppe et al., 1978; Sulston et al., 1983). The customary schematic view of the E lineage is shown on the left side of Figure 2. The right side of Figure 2 depicts the more realistic view of the intestinal lineage described by Leung et al (1999), which involves a series of cell divisions precisely oriented in three dimensions, together with short range cell migrations (intercalations) and dramatic intracellular polarizations. In the current section, we will consider only the cellular events that produce the intestine (Leung et al., 1999). The molecular mechanisms by which the E blastomere becomes specified will be described in a future Wormbook chapter by J. Rothman and M. Maduro and will also be summarized below in the section on transcriptional regulation.
|
Figure 2. Cell lineage of the C. elegans embryonic intestine. On the left is shown the conventional schematic lineage relating cell divisions and developmental time at 20°C. Anterior is to the left; posterior to the right; solid lines represent lineages on the left side of the animal; dotted lines represent lineages on the right side of the lineage. (Redrawn from Fukushige et al., 2005). Right side of the figure represents the disposition of the various E cells in three dimensions. Small dumb-bells connect sister cells. Red circles (with red arrows) at the bottom right of the figure represent E lineage nuclei that migrate towards the embryonic midline in the process of epithelial polarization. The right side of the Figure is redrawn from part of Figure 1A of Leung et al (1999); bilateral cell pairs in the intestine primordium are relabelled “a, b,..h” in order to retain the original nomenclature of the “ints” in the mature intestine, i.e. “int1, int2,..int9” (Sulston et al., 1983).
The first division of the E blastomere takes place on the surface of the embryo and produces an anterior daughter Ea and a posterior daughter Ep (see Figure 2). Ea and Ep then migrate into the embryo during gastrulation (see Gastrulation in C. elegans). The direction of the second cell division of the E lineage is left-right and provides the basis for the bilateral symmetry of the intestine; i.e., the left pair of cells, Eal and Epl, produce the left half of the intestine; the right pair of cells, Ear and Epr, produce the right half. The third division is basically anterior-posterior but the division axis for the anterior pair of cells is skewed in the dorsal-ventral plane (see Figure 2). After the fourth division, the intestinal primordium contains 16 cells lying in two tiers: ten cells in a dorsal tier and six cells in a ventral tier (Leung et al., 1999).
At the two-tiered sixteen cell stage (see Figure 2), cells in the intestinal primordium become polarized (Leung et al., 1999): intracellular components shift their relative positions, the cells change their shape and the future intestinal lumen begins to form between the two halves of the primordium. Nuclei of the 16E cells migrate towards the embryonic midline and cytoplasmic constituents (e.g., yolk granules and mitochondria) end up laterally. Despite the prominence of this nuclear migration, it is apparently not necessary: unc-83 mutants have a functional intestine but the nuclear migrations within the intestine primordium do not occur (Starr et al., 2001). The overall cellular polarization within the intestinal primordium appears to be an intrinsic property of E cells, since it can occur in isolated blastomeres (Leung et al., 1999).
Formation of the intestinal lumen begins with the appearance of small cavities at the midline between the left and right sets of cells. At the same time, small membrane-bounded vacuoles (0.3 to 0.5 microns in diameter) are detected close to the apposed central membranes (Leung et al., 1999). These vacuoles may be the same as the vacuoles described by (Roudier et al., 2005), which are endosomal in origin and contain the protein VPS-27 (vacuole protein sorting).
After the cells in the intestinal primordium have polarized, cells in the lower tier intercalate into the cells of the upper tier but cells do not cross the midline (Leung et al., 1999). As shown in Figure 2, the cell pair labelled “g” intercalates between cell pairs “c” and “d” and the cell pair “f” intercalates between cell pairs “a” and “b”. The anterior pair of cells (labelled “a” in Figure 2) then divide in a dorsal-ventral direction and the posterior pair of cells (labelled “h”) divide in an anterior-posterior direction, leading to the final arrangement of cells within the developing intestine: a series of nine rings referred to as “ints” (Sulston et al., 1983; White, 1988). The anterior ring (int1) has four cells and each of the eight more posterior rings (int2 through int9) has two cells. The basic form of the intestine at hatching is shown schematically at the bottom of Figure 2. A schematic view of the basic “int” unit of the C. elegans intestine is shown in Figure 3 below. The two semi-circular intestine cells (enterocytes) are attached to each other by apical junctions (Leung et al., 1999; see Epithelial junctions and attachments, and see below) and share the intestinal lumen between their opposed apical microvilli-containing surfaces. Enterocytes are also attached by apical junctions to their anterior and posterior neighbours.
Although the intestine is usually regarded as a simple tube, there are distinct patterning and morphogenetic events within the intestine in which certain ints behave differently than other ints. For example, the cells of int1 and int2 have shorter microvilli and form a collapsible conical reservoir at the intestine anterior; int5 cells interact intimately with the primordial germline; posterior ints are the seat of the defecation cycle, and; nuclear divisions and gene expression patterns can differ along the anterior-posterior intestine axis. The first example of intestinal patterning to be considered (and the best understood at the molecular level) is the intestinal twist that involves cells of int2, int3 and int4.
Midway through embryogenesis, at the point when the developing intestine has 20 cells, three of the “int” rings, int2, int3 and int4, undergo a clockwise rotation (as seen facing the embryo) of approximately 90° (Sulston et al., 1983); this rotation increases to approximately 180° by the time of hatching (Sulston and Horvitz, 1977) but may be variable (See WormAtlas). Presumably, the resulting twist allows the intestine to intertwine with the asymmetrically disposed hermaphrodite gonad. The cellular and molecular basis of this morphogenetic twist has been investigated by Hermann et al. (2000) and by Neves and Priess (2005), who have shown that the twist is the result of repeated application of the LIN-12/Notch pathway (see Notch signaling in the C. elegans embryo).
When the intestine primordium has four cells, the LIN-12 receptor can be detected on the surface of all four cells; no left/right asymmetry can be detected and this early phase of lin-12 gene expression appears to be controlled by the END-1 GATA factor. By the 8E cell stage, however, LIN-12 is markedly downregulated in cells of the left half of the primordium because particular MS-lineage cells expressing the LIN-12 ligand LAG-2 contact the intestine primordium on the left side, not on the right side. Thus, the intestine asymmetry can be traced to MS-lineage asymmetry, which in turn can be traced to the fundamental left/right asymmetry apparent at the six cell stage of the embyo (Wood, 1991; Wood et al., 1996). The LIN-12 asymmetry within the early intestine primordium sets the stage for a second phase in which the actual rotation takes place; this second phase also involves asymmetric LIN-12 receptor but now with the alternative ligand APX-1. Both of these interactions, the LAG-2 dependent imposition of LIN-12 asymmetry at the 4E stage and the subsequent APX-1 dependent process of cell rotation, involve the bHLH protein REF-1 as a direct downstream target of Notch signaling (see Notch signaling in the C. elegans embryo; Neves and Priess, 2005). The REF-1 effect is ascribed to a repression of downstream genes (in particular, the lin-12 gene itself in the first interaction) and involves the corepressor UNC-37/Groucho. The decision of which cells will rotate (i.e., what sets the anterior/posterior boundary of the twist) is controlled by the Wnt pathway implicated in the anterior-posterior patterning of gene expression, as will be described in the following section.
Fascinating questions of cell biology remain to be addressed. For example, how do these twisting cells break contact with their neighbours, rotate through this relatively precise angle, rearrange the complex three-dimensional scaffold of apical junctions with their neighbours, and yet still manage to exert the correct degree of rotational torque necessary to impart the helical form to the overall intestine?
A significant number of genes are expressed in specific non-uniform patterns along the anterior-posterior axis of the intestine: for example, genes associated with the defecation pacemaker are expressed towards the intestine posterior (Dal Santo et al., 1999), the ges-1ΔB transgene is expressed in the anterior int1 and int2 cells (Schroeder and McGhee, 1998) and the pho-1 acid phosphatase gene is expressed everywhere in the intestine except in int1 and int2 (Beh et al., 1991; Fukushige et al., 2005). Indeed, in situ hybridization (see Protocols for large scale in situ hybridization on C. elegans larvae, http://nematode.lab.nig.ac.jp/db2/index.php) reveals a number of clear examples where endogenous transcripts, not just reporter gene expression patterns, are expressed either in the intestine anterior, like ges-1ΔB, or everywhere but in the intestine anterior, like pho-1. It is well to keep in mind that reporter genes driven by incomplete promoters often express in the posterior intestine and, to a lesser extent, in the anterior intestine (Ruvinsky and Ruvkun, 2003; see Transcriptional regulation); thus, such expression patterns should be treated with caution.
The anterior-posterior patterning of gene expression is autonomous within the intestine lineage and reflects zygotic (i.e., non-maternal) application of the Wnt pathway (see Wnt signaling, Schroeder and McGhee, 1998), at least as worked out for the case of the ges-1ΔB transgene. At least the major components (WRM-1, LIT-1, and the downstream effector POP-1) of this pathway are the same molecules studied so intensely in the earlier P2-EMS contact that specifies the intestine. Overall, the ges-1ΔB transgene behaves as if its transcription is activated in the intestine anterior by high levels of the HMG protein POP-1 (Schroeder and McGhee, 1998). Conversely, expression of the pho-1 acid phosphatase gene behaves as if it is repressed in the intestine anterior by high levels of POP-1 (Fukushige et al., 2005). Because there are no obvious POP-1 binding sites in either promoter, POP-1 and the Wnt pathway may not act directly on the ges-1ΔB or pho-1 promoters but rather may exert their effects via fate changes earlier in the E lineage. As noted in the previous section, the same Wnt pathway also appears to limit the anterior-posterior extent of the intestinal twist (Hermann et al., 2000).
A further example of patterning within the intestine lineage is provided by the germ cell precursors extending lobes into the int5 cells of the intestine, apparently to obtain nutrition (Sulston et al., 1983). Edgar et al. (2001) found that the pal-1 homeobox gene (whose maternal phase specifies the fate of the C and D blastomeres (Hunter and Kenyon, 1996) is expressed in int5 beginning from approximately 350 cells until at least hatching and perhaps this is what makes int5 different from the other ints. However, int5-specific expression of pal-1 may not reflect patterning pathways within the intestine but rather regulatory control from within the primordial germ cells: the pie-1 gene appears to promote this germ cell-intestine interaction and pal-1 expression within the intestine might then be a downstream consequence (Tenenhaus et al., 2001).
At the time of hatching, the L1 intestine has 20 cells, each with a single diploid nucleus (Hedgecock and White, 1985). At the beginning of the L1 lethargus, intestinal nuclei replicate their DNA and divide in a spatially patterned but slightly variable manner: the six nuclei of int1 and int2 neither replicate their DNA nor divide; the four nuclei of int8 and int9 may or may not replicate and divide; the ten nuclei of int 3 to int7 always replicate and divide. As a result, the late L1 intestine retains 20 cells but now has 30 to 34 nuclei, all still diploid (Hedgecock and White, 1985). These nuclear divisions do not occur in null mutants of the heterochronic gene lin-14 (Hong et al., 2000) but the intestine appears to function nonetheless.
All intestinal nuclei endoreduplicate their DNA prior to the L1-to-L2 moult, whether or not they have just undergone one of the nuclear divisions described in the last paragraph. These endoreduplications occur without obvious chromosome condensations and are repeated before each of the three remaining moults, thereby producing the large 32C nuclei of the adult intestine (Hedgecock and White, 1985). Presumably, the basic C. elegans cell cycle machinery must be involved or must somehow be modified to produce these correctly-timed endoreduplications. For example, it has been shown that CKI-1 (a member of a family of cyclin dependent kinase inhibitors) is involved in limiting the number of cell divisions in the intestine. cki-1 knockouts, either by RNAi or by chromosomal deletions, lead to >30 nuclei, with the extra nuclei being produced by an extra round of division shortly before hatching (Hong et al., 1998; Kostic and Roy, 2002; Fukuyama et al., 2003). Simultaneous knockout of cki-1,2 and lin-35/Rb leads to an average of 84 intestinal nuclei (Boxem and van den Heuvel, 2001). The effects of cki-1 knockdowns are seen in many embryonic lineages, not just the intestine, and this holds true for mutations/RNAi in a number of other cell cycle control genes (see reviews by, Kipreos, 2005; Koreth and van den Heuvel, 2005; Boxem, 2006) or even in a calmodulin gene (Karabinos et al., 2003a). In contrast, two independent genetic screens identified mutations that affect only the intestine; both mutations result in an approximate doubling of both intestinal nuclei and intestinal cells, caused by an interpolated round of cell division at the 8E cell stage (Clucas et al., 2002; Kostic and Roy, 2002). In spite of this intestinal phenotype, both mutants are viable and fertile. The two mutations are both maternal dominant (or semi-dominant) gain-of-function, and are caused by single amino acid changes in adjacent residues in a CDC-25.1-like cell cycle phosphatase. Although these two specific gain-of-function alleles affect only the intestine, RNAi-induced loss of function of the same gene clearly affects many lineages and indeed the protein is widespread in the early embryo.
Finally, we recall the observation that expression of biochemical markers in the early E lineage depends upon a critical prior round of DNA synthesis occuring in the E cell, the clonal progenitor of the intestine (Edgar and McGhee, 1988). Perhaps it would be worthwhile to re-visit this observation in the light of the current detailed understanding of endoderm specification.
In forming the dauer larva, the alternative long-surviving third larval stage, the intestine becomes radially constricted and the lumenal circumference decreases by several-fold, to the point where the microvilli can become indistinct (Popham and Webster, 1979; Albert and Riddle, 1988). In the L2d stage, the L2 larval stage leading up to dauer formation, the intestinal cytoplasm can be packed with electron dense “cytosomes” (Popham and Webster, 1979; Albert and Riddle, 1988). How loss of intestinal volume and increase in density happens is not understood but autophagy could be involved (Melendez et al., 2003). It is also not known how the remodeling or the recovery pathways of the dauer intestine relate to the developmental pathway of the non-dauer intestine. The behaviour of particular mutants in daf-9 and daf-15 show that remodeling in the intestine can be separated from the remodeling of other tissues (Albert and Riddle, 1988).
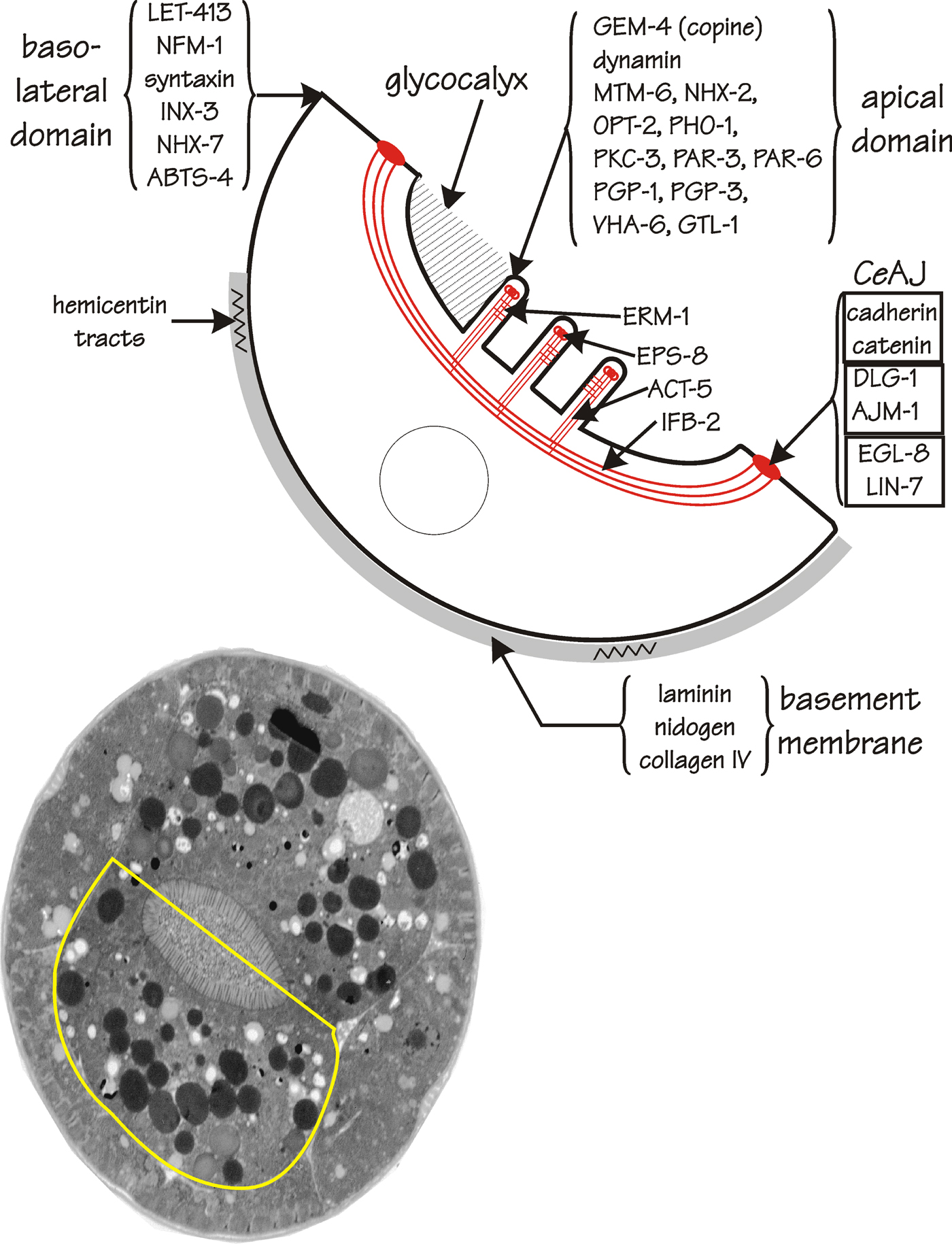
Figure 3 is a schematic representation of a C. elegans enterocyte, one of the pair of large, roughly centrosymmetric cells that surround the intestinal lumen and that constitute the basic unit of an “int” ring. The present section will emphasize the sub-cellular structures and molecular localizations within the enterocyte, leaving questions of function until later. The section is organized by cellular domain: (see section 4.1) the apical domain including brush border and terminal web; (see section 4.2) the basolateral domain including the basement membrane; (see section 4.3) the apical junctions joining one enterocyte to its partner and to adjacent ints within the overall intestinal structure, and; (see section 4.4) several of the intestine's organelles and inclusions. The spatial distribution of each protein discussed will be summarized on Figure 3, including proteins for which a role in intestine structure can reasonably be surmised (e.g., actin in microvilli) as well as proteins that were investigated for other reasons (often because of a role in neurons) but whose major site of expression then turned out to be in the intestine.
|
Figure 3. Structure of the C. elegans enterocyte. The transmission electron micrograph in the lower part of the figure is of a cross-section of a newly-hatched L1 larva (courtesy of Dr. J. Kormish and Mr. Wei Dong, University of Calgary). A single enterocyte, i.e., half of an “int”, is outlined in yellow. A schematic view of this enterocyte is shown at the top of the figure. Gene products (or other markers) that show differential localization to the apical or the basolateral domain are indicated, as described in more detail in the text.
The most prominent feature of the intestinal apical domain is the microvillar brush border (see Figure 3) and a number of genes integral to C. elegans microvilli have now been identified. The following summarizes the various points of interest.
Although it had previously been known that a YFP/GFP-tagged ACT-5 actin protein produced in the intestine would localize to the apical surface (Bossinger et al., 2004; Gobel et al., 2004). MacQueen et al. (2005) have now shown clearly that, at the ultrastructural level, ACT-5 resides within the microvilli. Knockouts of act-5 cause lethality shortly after hatching. The mutant intestinal cells retain normal polarity and have apical junctions but are completely devoid of microvilli. The intestinal lumen is now circular, rather than the usual squashed ellipse. Expression of the act-5 gene is limited to the intestine and several other cells with microvilli. Experiments to replace act-5 with other C. elegans actin genes were not successful, suggesting that ACT-5 performs a unique function in microvilli (MacQueen et al., 2005).
Proteins belonging to the ERM family (ezrin-radixin-moesin) are usually regarded as connecting the actin cytoskeleton to the plasma membrane and that is where ERM-1, the C. elegans homolog, is placed on Figure 3. Both antibody staining and GFP reporters place ERM-1 at or closely beneath the intestinal apical surface, certainly more apically disposed than are the apical junctions (Gobel et al., 2004; Segbert et al., 2004). RNAi to erm-1 is lethal, producing distorted intestines that appear to have adhesions across the lumen. These defects could arise from a failure in a general process of lumen formation (defects are also present in the excretory canal cell), from aberrant intercalation of cells during intestinal morphogenesis (see Figure 2 above) or from defects in the maturation of the apical junctions. As in the knockouts of act-5, the intestinal lumen of erm-1 knockouts is now circular rather than ellipsoidal and the microvilli are replaced by a disorganized tangle of fibrils, which may well be actin (Gobel et al., 2004).
The EPS-8 protein has been located by immunoelectron microscopy in the microvillar tips and, to a lesser extent, in the microvillar bases (Croce et al., 2004). Loss of EPS-8 function can be lethal, with obvious intestine defects: EPS-8 deficient microvilli appear less dense, less rigid and generally more irregular than wildtype microvilli. EPS-8 was also shown to have an actin-capping function in vitro and might function in the process of quality control, somehow regulating the remarkable uniformity of microvillar length.
The intermediate filament protein IFB-2 is the epitope reacting with the monoclonal antibody MH33 (Francis and Waterston, 1985; Karabinos et al., 2001; Bossinger et al., 2004) and is located within the “endotube” (Bossinger et al., 2004), a well-defined sub-apical layer within the terminal web of nematode intestines (Munn and Greenwood, 1984). This layer abuts directly onto the inter-enterocyte apical junctions, as seen by immunoelectron microscopy. Presumably, the function of the endotube is to provide structural strength and to anchor the overlying microvilli. The C. elegans genome contains 11 genes coding for intermediate filaments (Dodemont et al., 1990; Dodemont et al., 1994; Karabinos et al., 2001; Karabinos et al., 2002; Karabinos et al., 2003b; Karabinos et al., 2002; Karabinos et al., 2004) and six are expressed in the intestine (Karabinos et al., 2004); of these, three (in addition to IFB-2) are expressed exclusively in the intestine and produce proteins that, at least from light microscopy, are also located in the terminal web. These intestinal intermediate filaments probably function redundantly; intestinal malformations can be observed when several of the intermediate filament genes are simultaneously targeted by RNAi (Karabinos et al., 2004).
Electron micrographs show a fuzzy layer of material encasing the microvilli and extending slightly into the intestinal lumen (see Figure 3). Presumably this layer corresponds to the C. elegans glycocalyx or “peritrophic matrix” (Lehane, 1997), a complex, relatively unstructured collection of highly modified glycoproteins that serves several critical functions, including: (i) to protect the microvillar surfaces from mechanical injury or from pathogenic attack (see below); (ii) to serve as a filter allowing only digestion products to reach the absorptive surfaces, and; (iii) to provide a scaffold for presenting digestive enzymes to the contents of the intestinal lumen. The ultrastructure of this layer has been investigated in several nematodes, including C. elegans (Borgonie et al., 1995) and chemical stains reveal both heparin sulfate and dermatan sulfate in the region of the terminal web and brush border (Schimpf et al., 1999). Several lectins have been shown to bind to the edge of the intestinal lumen, some in an anterior-posteriorly patterned manner (Borgonie et al., 1994; Borgonie et al., 1997).
A number of further proteins show distinct apical localization within the intestine but may not play a primary role in enterocyte structure. Many of these proteins could be equally well discussed in later sections on digestion, metabolism, channels and transporters but are described here because what is presently known about their subcellular distribution appears more striking than what is known about their function:
PKC-3, a C. elegans atypical protein kinase C, is tightly localized to the apical surface of the intestine (Leung et al., 1999; Bossinger et al., 2001; Gobel et al., 2004; Van Furden et al., 2004). Together with the PAR-3 and PAR-6 proteins so critical in establishing the fundamental asymmetry of the early embryo (see Gastrulation in C. elegans), PKC-3 apical localization persists from mid-embryogenesis into adulthood and thus is unlikely to function only in the establishment of the intestinal apical-basal polarity (Wu et al., 1998). A protein (CKA1 for C kinase adaptor) was identified in a two-hybrid screen as interacting with PKC-3 and is thought to anchor PKC-3 to the cell membrane (Zhang et al., 2001). CKA1 is highly expressed in the intestine and shows significant sequence similarity to Drosophila and mouse Numb proteins.
The copine protein GEM-4 (a calcium dependent phosphatidylserine binding protein) is widespread in the embryo but after hatching, most GEM-4 is located in the intestinal brush border (Church and Lambie, 2003).
MTM-6 is one of the five myotubularins in C. elegans (phosphatidylinositol(3) phosphate phosphatases). MTM-6 is present almost exclusively in the intestine and is strongly localized to the apical membrane, consistent with roles in endocytosis and in controlling the level of inositol phosphates (Xue et al., 2003).
High levels of dynamin are associated with the lumenal surface of the intestine, consistent with high rates of endocytosis associated with retrieving food from the intestinal lumen (Labrousse et al., 1998).
The SEL-1 protein, originally identified through suppression of mutations in lin-12, can be detected in vesicles concentrated near the apical surface of intestinal cells (Grant and Greenwald, 1997).
Following heat shock, the small HSP16 proteins localize to discrete cytoplasmic structures bordering the intestinal lumen (Ding and Candido, 2000).
There must be many additional proteins attached to the intestine lumenal surface but only a small fraction have been identified: e.g., the PHO-1 intestinal acid phosphatase GPI-anchored to the intestine lumenal surface (Beh et al., 1991; Fukushige et al., 2005), the OPT-2/PEP-2 peptide transporter (Nehrke, 2003; Meissner et al., 2004), the VHA-6 vacuolar ATPase subunit (Oka et al., 2001) and the NHX-2 sodium-hydrogen exchanger (Nehrke and Melvin, 2002; Nehrke, 2003). Several of these proteins will be discussed in later sections.
The basement membrane surrounding the intestine (see Figure 3) has three of the four major constitutents found in other C. elegans basement membranes: laminin α and β chains (see Basement membranes; Huang et al., 2003; Kao et al., 2006), nidogen/entactin (Kang and Kramer, 2000) and type IV collagen (Graham et al., 1997), all detected on the surface of the intestine beginning in early morphogenesis phase. Interestingly, type IV collagen is not synthesized in the intestine but rather in muscles and in somatic gonads (Graham et al., 1997), from which the product must be able to diffuse through the pseudocoelom to assemble into the basement membrane covering the intestine as well as the pharynx. In contrast, perlecan assembly appears to be cell-autonomous (Moerman et al., 1996) but is not found in the basement membrane of the intestine (Norman and Moerman, 2000). Longitudinal tracts of hemicentin, a large member of the immunoglobulin superfamily, are produced by muscles and gonads and apparently mechanically anchor the intestine, probably via the intestinal basement membrane, to the regions between muscles and hypodermis (Vogel and Hedgecock, 2001).
The following proteins have been associated with the basolateral domain of intestine cells:
LET-413, the homolog of Drosophila scribble (Legouis et al., 2000; Legouis et al., 2003; Bossinger et al., 2004; Segbert et al., 2004) is distributed throughout the basolateral domain. Depletion of LET-413 causes the apical junctions to become extended and discontinuous; more dramatically, apical markers such as the entire terminal web, now extend to the basolateral surface (Legouis et al., 2000; McMahon et al., 2001; Bossinger et al., 2004). Thus LET-413 appears to act as a fence to confine the apical domain but has also been proposed to function in polarized protein trafficking (Legouis et al., 2000).
NFM-1, the C. elegans homolog of neurofibrularin, is located throughout the basolateral domain, a distribution more-or-less complementary to that of its relative ERM-1 (Gobel et al., 2004).
Syntaxin is expressed strongly in neurons but is also detected on the basolateral surface of the intestine (Saifee et al., 1998).
INX-3 is a gap junction channel protein, widely expressed in the embryo. At the comma stage, INX-3 becomes strongly (if transiently) concentrated to the membranes between intestine cells (Starich et al., 2003).
Discrete electron dense regions can be detected just beneath the apical, lumenal surface of the intestinal cells, joining each cell to its neighbour (Legouis et al., 2000). The nomenclature of these structures has changed over the years but they are now generally referred to as “apical junctions”. The C. elegans apical junctions are regarded as the counterpart of the bipartite cell junctions seen in Drosophila epithelia (zonula adherens and septate junctions) and the tripartite structures seen in vertebrate epithelia (tight junctions, zonula adherens and desmosomes). Indeed, many of the proteins found in fly and vertebrate cell junctions have clear homologs in C. elegans and this is how many of the C. elegans components were identified. However, the function of each component and its position in the assembly pathway in C. elegans epithelia can differ markedly from other systems. As particular examples, depletion of cadherins, catenins and proteins such as crumbs (CRB-1) has disasterous consequences for fly epithelia but relatively minor consequences in the C. elegans intestine (Costa et al., 1998; Bossinger et al., 2001). One particular view of the relative positioning of proteins and protein complexes within the apical junctions, is shown in Figure 3. The WormBook chapter by Labouesse (see Epithelial junctions and attachments) provides a much more detailed discussion of the important topic of apical-basal polarity in C. elegans epithelia, including the intestine.
Two other interesting genes are highly expressed in the intestine, with the encoded proteins apparently located in the apical junctions:
EGL-8 is a phospholipaseCß with a documented function in the nervous system (Miller et al., 1999) and may play a role in oscillating Ca++ signaling (see below).
LIN-7 is a PDZ-containing protein well studied for its interactions with the LET-23 receptor tyrosine kinase in vulva induction (Simske et al., 1996).
The well-known birefringent “gut granules” that have served so well as markers of intestinal differentiation have been shown to be lysosome-related organelles (Clokey and Jacobson, 1986; Hermann et al., 2005). The evidence derives from staining with a number of lysosome-specific dyes and protein markers, as well as showing that gut granule formation is prevented by ablating genes whose homologs are known to function in lysosome biogenesis in other experimental systems. (Hermann et al., 2005) have screened for a gut granule loss (Glo) phenotype in order to identify genes involved in lysosome formation. In some of these Glo mutants, the birefringent material is no longer retained within intestinal cells but rather is mis-routed into the intestinal lumen. GLO-1 is a RAB GTPase expressed only in the intestine and is indeed found to be associated with the granules. The identity of other genes in this pathway have suggested that gut granules may be similar to pigment granules in Drosophila and melanosomes in humans (Hermann et al., 2005).
Several other proteins have been associated with gut granules or with related intestinal organelles:
The VHA-11 protein is a subunit of a vacuolar ATPase and appears in embryonic intestinal cells at the comma stage in punctae that could be gut granules. It is proposed that VHA-11-containing vacuolar ATPase contributes to acidification of cellular compartments, crucial to intracellular sorting and endocytosis (Oka and Futai, 2000).
LMP-1 is the C. elegans homolog of mammalian “lysosome-associated-membrane-protein”. Beginning at the lima bean stage of early embryogenesis, LMP-1 can be detected on the periphery of a class of intestinal granules that also seem to be good candidates for “gut granules” (Kostich et al., 2000). In the early embryo, LMP-1 protein is more widespread and is associated with the cell membrane and perhaps earlier stages in the endosomal compartment (Hermann et al., 2005). A knockout of the lmp-1 gene has little effect on the general health of the animal but one particular size of intestine granules is now missing (Kostich et al., 2000). Chen et al. (2006) have described a number of new protein markers that are beginning to reveal the true diversity of vesicles in the intestine.
C. elegans, like mammals, appears to have two sites of β-oxidation of fatty acids: mitochondria and peroxisomes (Gurvitz et al., 2000) and most C. elegans peroxisomes are found in the intestine, as shown by immunostaining for catalase (Togo et al., 2000) and by electron microscopy (Yokota et al., 2002). The use of RNAi to inhibit peroxisome biogenesis leads to complete L1 arrest, much like starvation (Thieringer et al., 2003).
The C. elegans pex-2 and pex-6 peroxin genes are expressed mainly in the intestine, as shown by in situ hybridization (Ghenea et al., 2001). In other systems, peroxins are associated with peroxisome biogenesis.
The enzyme Type II 3-oxoacyl-coA thiolase (also called sterol carrier protein) is expressed largely in the intestine and is located within the peroxisome matrix; levels of this enzyme are induced several-fold by the peroxisome proliferator drug clofibrate (Maebuchi et al., 1999).
It is difficult to disagree with the lament that “unfortunately, the biology of digestion (in C. elegans) represents something of a blind spot in this otherwise remarkably well-characterised organism” (Walker et al., 2005). However, digestion and metabolism have been studied in other nematodes (Behm, 2002; Munn and Munn, 2002) and there is increasing understanding of these processes in C. elegans as well. For example, analysis of a SAGE library prepared from dissected adult intestines identifies transcripts from over four thousand genes (McGhee et al., 2007) and this list of genes must hold many keys to understanding digestion, metabolism and other aspects of intestinal physiology. In this section, we will highlight genes that are expressed highly, often exclusively, in the intestine and for which a functional role in digestion and metabolism is either known or plausible.
The general steps by which the animal digests its bacterial food can be surmised and are fully supported by analysis of the most highly expressed, most intestine-restricted set of genes identified in our SAGE analysis (McGhee et al., 2007). Presumably, the first act of digestion is physical damage to the bacteria inflicted by the pharyngeal grinder (Avery and Thomas, 1997). A small number of lysozymes (Mallo et al., 2002; McGhee et al., 2007) are highly expressed in the intestine, are probably secreted and are the likely perpetrators of the next step in digestion, degradation of the bacterial cell wall. Several saposins/amoebapores are also highly expressed in the intestine and some appear to be secreted; presumably these small pore-forming proteins are able to puncture the bacterial membranes to release contents that can now be degraded by lumenal hydrolases (Banyai and Patthy, 1998).
What is known about the general properties of the lumenal environment in which digestive hydrolases must act? For example, what is the lumenal pH? Most protease activity in crude extracts likely derives from intestinal proteases (see below) and the pH optimum of this overall protease activity is approximately 5 (Sarkis et al., 1988; Geier et al., 1999). The pH optimum of the PHO-1 acid phosphatase, which is GPI-anchored in the intestinal lumen, is approximately 4 (Beh et al., 1991). These two observations suggest that the intestinal lumen must be mildly acidic. Considering that roughly half of the intestinal contents are ejected with each defecation cycle of less than one minute (Avery and Shtonda, 2003; Ghafouri and McGhee, 2007) and considering that C. elegans can live in highly buffered media, the acidification capacity of the intestine must be enormous. Indeed, several vacuolar ATPase subunits are highly expressed in the intestine and, in particular, the VHA-6 protein is localized to the apical surface of the intestine, where it is suggested to acidify the intestinal lumen as part of the mechanism of nutrient uptake (Oka et al., 2001).
Proteases/peptidases appear to be more prominent than any other class of lumenal digestive hydrolases. For example, 18 of the top 80 genes expressed largely (or exclusively) in the adult intestine are proteases/peptidases and five of these are aspartic proteases (McGhee et al., 2007). Indeed, >90% of the protease activity in crude C. elegans extracts can be inhibited by pepstatin, a specific inhibitor of aspartic proteases (Geier et al., 1999). Probably the most thoroughly characterized digestive enzyme in C. elegans is the aspartic protease ASP-1, which was shown to be expressed exclusively in the intestine; ASP-1 appears late in embryogenesis, reaches peak levels during the first two larval stages and declines thereafter (Tcherepanova et al., 2000). Antibody staining indicated that most ASP-1 protein is in the intestinal lumen, although some protein could also be detected in lysosome-like cytoplasmic punctae (Tcherepanova et al., 2000). Other peptidases, some apparently quite specialized, are also highly expressed in the intestine: the PAM-1 puromycin sensitive aminopeptidase (Brooks et al., 2003), the APP-1 aminopeptidase (Laurent et al., 2001) and the LAP-1 leucine aminopeptidase (Joshua, 2001).
Nematodes rely heavily on lipids for energy storage and energy metabolism (Behm, 2002) and the intestine appears to be a major site of these biochemical activities. Lipases are highly represented on the SAGE list of intestinally-expressed genes and a number of these are likely to be secreted and function in lumenal digestion of lipids (McGhee et al., 2007). It was mentioned in the previous section that peroxisomes are one of the two sites of fatty acid oxidation in C. elegans and most peroxisomes are in the intestine. A number of enzymes involved in lipid metabolism are highly expressed in the intestine: for example, cln-3.1 is homologous to a human gene involved in the degenerative disorder neuronal ceroid lipofuscinosis (de Voer et al., 2005) and the spl-1 gene encodes sphingosine phosphate lyase (Mendel et al., 2003; Oskouian et al., 2005). The ELO-2, ELO-5 and ELO-6 enzymes have fatty acid elongation activities and are involved in the synthesis of polyunsaturated and branched fatty acids (Kniazeva et al., 2003; Kniazeva et al., 2004). C. elegans has three Δ-9-desaturase enzymes, expressed primarily (fat-5 and fat-6) or exlusively (fat-7) in the intestine (Brock et al., 2006); single knockouts have little phenotype but the triple knockout is lethal, demonstrating an overall requirement of C. elegans for adequate levels of unsaturated fatty acids (Brock et al., 2006). Finally, the GES-1 nonspecific carboxylesterase resides in the intestinal endoplasmic reticulum (McGhee, 1987; Kennedy et al., 1993) and could function in lipid metabolism and/or detoxification.
The following list contains a brief description of other genes that are all expressed highly and often exclusively in the intestine. The list provides a hint of the enormous metabolic diversity of the C. elegans intestine:
The NUC-1 nuclease was originally identified because mutants retained bacterial DNA undigested within the intestine (Sulston, 1976); (also, unpublished results of J.E. Sulston and of P. Babu, cited in, Hevelone and Hartman, 1988). NUC-1 also functions in the removal of apoptotic corpses (Wu et al., 2000).
The TRX-1 thioredoxin expression in the intestine increases with starvation and aging; it is suggested that TRX-1 provides protection against oxidative damage associated with the metabolically active intestine (Jee et al., 2005).
cdd-1 and cdd-2 encode two cytidine deaminases highly expressed in the intestine. Similar enzymes are usually associated with pyrimidine salvage pathways but the two C. elegans proteins CDD-1 and CDD-2 have been shown also to bind RNA, thereby raising the possibility of an RNA editing pathway in the C. elegans intestine, perhaps similar to the editing of apolipoprotein B mRNA in the mammalian intestine (Thompson et al., 2002).
At least three of the 21 C. elegans Skp1-related genes (involved in ubiquitin-mediated protein turnover) are expressed highly in the intestine, suggesting a dedicated pathway to control the degradation of intestinal proteins (Yamanaka et al., 2002).
ftn-1, one of the two C. elegans ferritin-encoding genes, is expressed exclusively in the intestine, from embryo to adult, and is induced at the transcriptional level by exposure to iron (Kim et al., 2004).
The three enzymes ornithine decarboxylase, S-adenosylmethionine decarboxylase and spermidine synthase are expressed primarily in the intestine (Luersen et al., 2004), suggesting that the intestine provides the animal's supply of polyamines.
Calreticulin (CRT-1) is a calcium-binding molecular chaperone expressed predominantly in the intestine, beginning from mid-embryogenesis (after an apparently ubiquitous early maternal phase). CRT-1 may be involved in the intestinal stress response; in addition, crt-1 genetically interacts with the itr-1 IP3 receptor and shows synergistic effects both on the defecation cycle and on brood size (Park et al., 2001).
Three p38 homologs (members of the MAPK superfamily) are arranged as a three-member operon, are expressed apparently exclusively in the intestine (Berman et al., 2001) and may be involved in the intestinal stress response.
The PHO-1 intestinal acid phosphatase is highly similar to lysosomal acid phosphatases in other species but most of the enzymatic activity resides at the edge of the intestinal lumen (Beh et al., 1991), retained by a GPI anchor (Fukushige et al., 2005). pho-1 knockouts are maternal effect lethal, suggesting that mothers require PHO-1 to process some nutrient that will then be supplied to the next generation embryos (Fukushige et al., 2005).
The gob-1 gene was identified because mutant larvae have a lethally obstructed intestine; GOB-1 turns out to be the enzyme trehalose-6-phosphate phosphatase and the lethality turns out to be caused, not by the absence of trehalose, but by a buildup of the intermediate trehalose-6-phosphate; a similar toxicity is seen in corresponding yeast mutants, emphasizing both the conservation and the expected future complexities of metabolic regulation (Kormish and McGhee, 2005).
The FUT-2 fucosyltransferase is expressed exclusively in the intestine, beginning in mid-embryogenesis; enzymes with related activity are found in mammalian intestines and are involved in sustaining bacterial flora (Zheng et al., 2002).
GLY-3 and GLY-5 are N-acetylgalactosamine transferases that produce mucin type linkages (Hagen et al., 2001).
GLY-14 and GLY-19 are N-acetylglucosaminyl transferases expressed exclusively in the intestine, from early larval stages through to adulthood (Chen et al., 1999; Warren et al., 2001).
Deletion of the Glc-NAC transferase gene causes intestinal glycogen stores to increase several fold and intestinal fat stores to decrease several fold (Hanover et al., 2005); the homologous enzyme in other systems may be involved in nutrient sensing, perhaps suggesting wider and more complex metabolic control in C. elegans as well.
Combined with the C. elegans strengths of classical genetics and RNAi, recent technical advances (e.g., the ability to image intracellular fluorescent dyes, fluorescent substrates and environmentally responsive GFP variants and the ability to directly investigate the electrophysiology of dissected intestines or intestinal cells) hold great promise for understanding nutrient transport, ionic homeostasis and complex behaviours such as defecation.
The C. elegans genome contains nine genes encoding sodium-hydrogen exchangers and three of the genes are expressed exclusively in the intestine (Nehrke and Melvin, 2002). Translational fusions with GFP show that NHX-2 is located on the apical membrane, NHX-7 is located in the basolateral membrane and NHX-6 appears to be a mixture, basolateral at the ends of the intestine and apical in the central. Perfusion experiments, combined with intestine-specific expression of pH-sensitive GFP variants, were used to investigate how C. elegans controls the intracellular pH of the intestine and, more particularly, how the NHX-2 exchanger compensates for the acidification caused by uptake of food-derived peptides (Nehrke, 2003). The OPT-2/PEP-2 protein is the major C. elegans dipeptide transporter and is also located on the lumenal surface of the intestine (Nehrke, 2003; Meissner et al., 2004).
The intestine is the most likely site for the receptor/transporter responsible for uptake of double-stranded RNA from the environment when dsRNA mediated interference (RNAi) is performed by feeding or soaking. Several candidate genes have been described. SID-1 is a multi-transmembrane protein located at the periphery of most C. elegans cells but appears especially enriched in cells, such as the intestine, exposed to the environment (Winston et al., 2002a). SID-1 functions cell autonomously, is necessary for systemic RNAi and can confer dsRNA uptake capabilities on heterologous cells (Winston et al., 2002b; Feinberg and Hunter, 2003). SID-2 is less well characterized but is also required for effective RNAi following feeding or soaking and is strongly expressed in the intestine (personal communication from Craig Hunter, cited in Britton and Murray (2006)). The haf-6 gene encodes a half molecule ABC transporter expressed predominantly in the intestine and germline; haf-6 mutants are especially defective in RNAi administered by feeding (Sundaram et al., 2006). A further eight ABC transporter genes (of the approximately 60 such genes in C. elegans) show effects in RNAi uptake or spreading and several of these are expressed in the intestine (Sundaram et al., 2006).
Brief descriptions of other intestinal transporters are provided in the following list:
ABTS-4 is an intestinal anion bicarbonate transporter and appears localized to the basolateral surface, at least as revealed by GFP translational fusions (Sherman et al., 2005).
Several intestinally-expressed sulfate permeases also appear localized to the basolateral membrane (Sherman et al., 2005).
The NAC-2 sodium-coupled citrate transporter (similar to Drosophila “I'm not dead yet”) is expressed throughout the intestine and is suggested to take up citrate and other TCA-cycle intermediates from the intestinal lumen for energy production (Fei et al., 2004). The NAC-1 and NAC-3 sodium-dicarboxylate anion transporters are also expressed strongly and exclusively in the intestine (Fei et al., 2003).
Two nucleoside transporters ENT-1 and ENT-2 are also expressed in the intestine (and pharynx; Appleford et al., 2004).
Two of the four TRPM-family channels in C. elegans, GTL-1 and GON-2, are expressed predominantly in the intestine; translational fusions with GFP show that the GTL-1 protein accumulates at the intestinal apical surface. Both GON-2 and GTL-1 play major roles in Mg++ ion transport and homeostasis in the intestine (Teramoto et al., 2005).
The C. elegans defecation motor program coordinates events that occur in the intestine, in muscles (especially enteric muscles) and in a small number of neurons (Avery and Thomas, 1997). A recent review (Branicky and Hekimi, 2006) lists >50 genes acting in one phase or another of the pathway and approximately 20 of these genes act in the intestine.
An important and striking finding was that the basic 45-50 second interval of the defecation cycle is associated with cyclic fluctuation in calcium ion levels in the posterior intestine; moreover, these calcium fluctuations reflect the behaviour of the inositol trisphosphate receptor ITR-1 (Dal Santo et al., 1999). Recently, powerful new techniques (e.g., electrophysiology, loading with calcium sensitive dyes, expression of calcium-sensitive GFP variants, combined with RNAi) have been applied to understand the molecular basis of these calcium fluctuations. Distinct calcium conductances have been identified by patch-clamping isolated intestinal cells (Estevez et al., 2003; Estevez and Strange, 2005). Dissected intestines (as long as they remain attached at the posterior to the carcass) are capable of sustained rhythmic oscillations in calcium ion levels (Espelt et al., 2005; Teramoto and Iwasaki, 2006). The rise in calcium ion levels closely corresponds to the pBoc phase of the defecation motor program (Espelt et al., 2005; Teramoto and Iwasaki, 2006). Waves of fluorescence reflecting transient increases in calcium ions can be observed to propagate from the posterior to the anterior intestine (Espelt et al., 2005; Teramoto and Iwasaki, 2006); the oscillations in fluorescence appear tightly focused on the apical region of the intestinal cell (Espelt et al., 2005). An example of this remarkable behaviour is shown in Movie 1 (Teramoto and Iwasaki, 2006). Blocking propagation of the calcium wave half-way down the intestine halts the defecation motor program, suggesting that the calcium wave is responsible for triggering the later muscle and neuronal phases (Teramoto and Iwasaki, 2006). Espelt et al. (2005) have extended such assays to show that only two of the six phospholipase C genes in C. elegans, egl-8 and plc-3, influence the defecation behaviour, either cycle time or cycle regularity. It was mentioned earlier that egl-8 is expressed highly in the intestine, with the EGL-8 enzyme concentrated near the apical junctions (Miller et al., 1999); plc-3 is also expressed in the intestine (Espelt et al., 2005). Intestine-specific RNAi was used to demonstrate that egl-8, plc-3 and itr-1 do indeed function in the intestine. It was further suggested that PLC-3 is responsible for generating the inositol triphosphates that control ITR-1 activity; in contrast, EGL-8 seems to function in a separate pathway to maintain the rhythmic oscillations in calcium ion (Espelt et al., 2005).
Movie 1. Intercellular Ca2+-wave propagation in the C. elegans intestine. Sequential images of the intact intestine in a living animal using the Ca2+-sensitive-fluorescent cameleon system. After Ca2+ spikes at the intestine posterior, a Ca2+ wave propagates from the posterior to the intestine anterior. The movie and its description have been reproduced from Teramoto and Iwasaki (2006) with permission from Elsevier.
flr-1 mutants were originally isolated on the basis of resistance to fluoride (Katsura et al., 1994) but turned out to have a short defecation interval. FLR-1 is a degenerin/epithelial sodium channel type of protein, expressed only in the intestine and possibly concentrated at apical membranes (Take-Uchi et al., 1998). The flr-4 gene encodes an apparently membrane-associated kinase that is expressed in the intestine (+ pharynx and several neurons) beginning from the comma stage; only intestinal expression is necessary for mutant rescue. GFP translational fusions to FLR-4 appear localized to the enterocyte membranes, especially the lateral membranes (Take-Uchi et al., 2005). Proteins binding to the ITR-1 protein in a yeast two-hybrid assay have also been identified in the intestine (and other tissues; Walker et al., 2004). All of the above genes are candidates for participating in signals that originate from the oscillating calcium levels in the posterior intestine and that result in contraction of the posterior body wall muscles (the pBoc phase of defecation). Cycle times are influenced by additional intestinally-expressed genes, for example, genes that influence fatty acid composition (Branicky and Hekimi, 2006).
The C. elegans intestine becomes active in endocytosis at least by the 8E cell stage, when there is transfer of yolk into the intestine primordium from the remainder of the embryo (Bossinger and Schierenberg, 1996; Yu et al., 2006). Once the animal hatches and begins to feed, nutrient uptake and trafficking must become an even larger fraction of intestinal activity, one that is just beginning to be investigated. Digestive and protective molecules must be secreted into the intestinal lumen and displayed on the apical plasma membranes. Nutrients must be taken up, either by endocytosis or by dedicated transporters. Processed food must be passed on to the rest of the animal through basolateral domains, either as building blocks or as synthesized products such as the vitellogenins. Similarly, there must be a variety of molecules, ranging from waste products to signals, passing in the opposite direction, back across the intestine basal surface from the rest of the animal. Thus one would expect that the C. elegans intestine is a hotbed of endocytosis, exocytosis and trafficking in general. Consistent with such a view, the intestine cell cytoplasm is filled with rough endoplasmic reticulum but smooth ER is either rare or absent (Rolls et al., 2002).
The process of endocytosis has been most productively studied in oocytes and in coelomocytes (Fares and Grant, 2002) but the identified genes are probably common to most C. elegans cells and indeed are now beginning to be identified in the intestine (Sato et al., 2005; Chen et al., 2006). See also the WormBook chapter “Intracellular trafficking” which discusses the general topic of intracellular trafficking in C. elegans in the detail it deserves.
Transport of a toxic derivative of linolenic acid from the intestine to the gonad may involve proteins such as NRF-5, NRF-6 and NDG-4 (Watts and Browse, 2006). The nrf genes were originally identified because of an altered response to Prozac (fluoxetine) (Choy and Thomas, 1999) and one class of these genes is proposed to be involved in transport of hydrophobic molecules from the intestine to the rest of the animal (Choy et al., 2006). In particular, NRF-5 is a secreted lipid binding protein similar to the cholesterol ester binding protein of higher organisms (Choy et al., 2006).
Considering the C. elegans habitat and dietary practices, it is not surprising that the intestine is a major site of response to toxins and chemical stressors in the environment. The C. elegans genome has two metallothionein genes (mtl-1 and mtl-2), both encoding proteins that are cysteine-rich but otherwise not closely related to metallothioneins described in other organisms (Slice et al., 1990; Freedman et al., 1993). Both genes are induced at the transcriptional level by exposure to low levels of cadmium (in the dose range of 1-100 micromolar), with a response time on the order of hours. The mtl-2 gene is expressed exclusively in the intestine, from L1 larvae through to adults, and either metal exposure or heat shock are required for detectable expression. The mtl-1 gene is also inducible in the intestine but has a minor constitutive component in several cells of the pharynx (Freedman et al., 1993). Gene knockouts show that MTL-1 and MTL-2 do indeed protect against cadmium toxicity (Swain et al., 2004).
There are other pathways besides metallothioneins that protect C. elegans against metal toxicity and several dozen genes responding to cadmium exposure have been identified (Liao and Freedman, 1998). One gene in particular (called cdr-1 for cadmium responsive gene) has been studied in greater detail (Liao et al., 2002). Like mtl-1 and mtl-2, cdr-1 is induced at the transcriptional level with a response time of hours but induction is highly specific for cadmium; expression of cdr-1 is exclusively in the intestine in all stages post-hatching. The CDR-1 protein is highly hydrophobic, appears novel and has been located in intestinal lysosomes (Liao et al., 2002). RNAi-induced loss-of-function shows that CDR-1 does indeed protect against cadmium toxicity. A series of related genes (cdr-2 through cdr-7) have now been identified (Dong et al., 2005) but it is not known whether these genes are also expressed in the intestine.
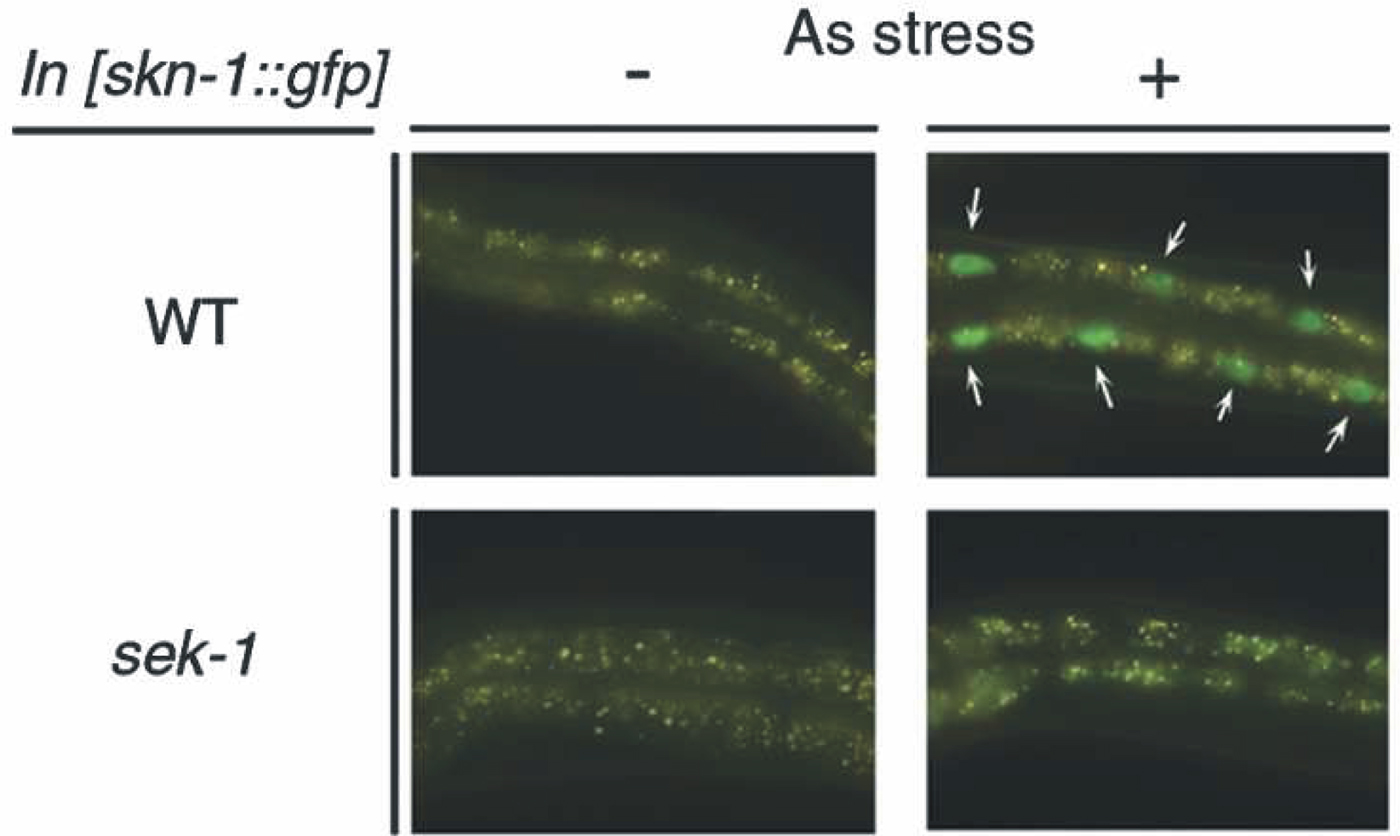
The maternally-provided SKN-1 transcription factor has been well studied for its role in specification of EMS cell fate in the early embryo but SKN-1 has long been suspected of having an additional role in intestinal differentiation and/or intestinal function (Bowerman et al., 1992; Bowerman et al., 1993). The proposed somatic function of SKN-1 is to regulate a bank of genes, possibly all intestinal, that are involved in “phase II” detoxification and that provide protection against reactive oxygen species and other environmental toxins (An and Blackwell, 2003). One example of such a gene is gcs-1, encoding the enzyme glutamylcysteine synthetase involved in the production of glutathione. A gcs-1::GFP reporter construct is expressed in the intestine, pharynx and ASI neurons; intestinal expression is induced by heat or by oxidative stress and is abolished either by mutating a SKN-1 binding site in the gcs-1 promoter or by abolishing SKN-1 activity (An and Blackwell, 2003). Thermal or oxidative stress causes a SKN-1::GFP reporter protein to relocate from intestinal cytoplasm to intestinal nuclei within minutes (see Figure 4). The distribution of SKN-1 protein between nucleus and cytoplasm reflects competition between two kinases: glycogen synthase kinase-3 and p38 MAP kinase (An et al., 2005; Inoue et al., 2005). Consistent with a function of SKN-1 in regulating intestinal detoxification genes, skn-1 mutants show shorter lifespans and are more sensitive to agents such as paraquat (An and Blackwell, 2003).
|
Figure 4. The C. elegans p38 MAPK pathway regulates the nuclear localization of SKN-1 in response to oxidative stress. Animals containing an integrated transgenic array of a SKN-1::GFP fusion reporter were exposed (+) or not exposed (-) to oxidative stress (5 mM arsenite for 1 hour). The nuclear relocation of the SKN-1 protein depends on the MAPK pathway, because it does not occur in mutants of the sek-1 gene (encoding a MAPKinase Kinase). Reproduced from Figure 2 of Inoue et al. (2005) with permission from CSHL Press.
The C. elegans genome contains approximately 80 genes encoding cytochrome P450 proteins, among whose roles is the metabolism of “xenobiotics”. A number of C. elegans P450 genes respond to compounds that induce mammalian cytochrome P450 genes (Menzel et al., 2001). Transcription of one gene in particular, CYP35A2, is strongly induced by exposure to ß-naphthoflavone and expression is exclusively in the intestine.
The four C. elegans P-glycoproteins (PGP-1 to PGP-4) belong to a family of membrane transporters containing an ATP-binding cassette. P-glycoproteins are often found on absorptive/excretory epithelial surfaces in higher organisms, where they are involved in the excretion of large hydrophobic toxins encountered in the diet or, for humans, encountered during chemotherapy. The pgp-1 and pgp-3 genes are expressed heavily in the C. elegans intestine throughout the lifespan (Lincke et al., 1993); antibody staining reveals that the proteins are located in the apical membranes facing the intestinal lumen (Broeks et al., 1995). pgp-3 is also expressed highly in the excretory cell. A deletion of the pgp-3 gene increases sensitivity to colchicines and to chloroquine (Broeks et al., 1995). The pgp genes, together with the related mdr genes, also contribute to protection against heavy metals (Broeks et al., 1996).
During the past decade, an entirely new field has developed in which C. elegans is used as a host to study the effects of pathogenic micro-organisms. The response of C. elegans to such pathogens is varied, ranging from no apparent effect up to killing within hours (in this case, usually because of a secreted toxin; see below). Probably the most common observation is that pathogenic bacteria are able to visibly and persistently colonize the intestine; indeed, a summary of responses of C. elegans to a series of pathogenic microbes (28 bacteria and two fungi) lists intestine as the main target (Schulenburg et al., 2004). This infectious interaction between microbes and C. elegans is beyond the scope of the present chapter and here we focus only on the best understood example of how the C. elegans intestine responds to a secreted bacterial toxin.
Aroian and coworkers have used C. elegans to study interactions with a set of crystal toxins produced by Bacillus thuringiensis (of agricultural fame) and the intestine appears to be the major target; see also the earlier work of Borgonie and coworkers (Borgonie et al., 1995; Borgonie et al., 1996). Exposure of C. elegans to purified toxin causes the intestine to become vacuolated within hours, followed ultimately by death (Marroquin et al., 2000). A series of BT-toxin-resistant mutants (bre-1 to bre-5) were isolated and the corresponding genes subsequently cloned. The major conclusion is that toxin resistance is associated with a series of glycosyltransferases, some of which are highly related to Drosophila genes with apparently similar function (Marroquin et al., 2000; Griffitts et al., 2001; Griffitts et al., 2003; Griffitts et al., 2005). The “bre” genes thus appear to form a conserved glycosylation pathway, the end product of which forms the toxin receptor or at least part of the receptor. Indeed, the intestines of resistant mutants no longer take up labeled toxin and expression of the appropriate bre gene only in the intestine is sufficient to restore sensitivity (Griffitts et al., 2003). A major (if not the major) receptor has now been shown to be a glycolipid, of a type common to invertebrates but not found in vertebrates (Griffitts et al., 2005); it is expected that this glycolipid forms part of the intestine lumenal membranes. Finally, microarrays have been used to identify C. elegans genes differentially regulated by exposure to Bt toxins; such genes include mitogen-activated protein kinases, which were shown to be important for normal toxin resistance (Huffman et al., 2004). Overall, these studies provide an encouraging example how C. elegans can be used to investigate problems of immense importance for world economics and public health.
There are a number of reasons to suspect that the adult intestine could play a central role in aging and lifespan. Dietary restriction can increase lifespan in C. elegans as in other organisms (Walker et al., 2005; Braeckman et al., 2006) and the intestine must be involved at some level in this phenomenon. Lipid accumulation responds to the daf-2/daf-16 signalling pathway and the intestine is a major site of lipid storage (Ashrafi et al., 2003). Expression of daf-16 only in the adult intestine is able to reconstitute much of the daf-16-dependent lifespan-extension in wildtype animals and essentially all of the daf-16-dependent lifespan extension in animals lacking a germline (Libina et al., 2003). Moreover, in animals lacking a germline (and which are consequently long-lived), DAF-16 protein accumulates in intestinal nuclei (Lin et al., 2001). Finally, reasonable connections can be advanced between aging, lifespan, stress responses (Hsu et al., 2003; Lamitina and Strange, 2005) and the fact that, as discussed in an earlier section, many of C. elegans stress responses are focused in the intestine.
Microarray analyses have identified genes whose expression is influenced by daf-2 or daf-16 mutations (McElwee et al., 2003; Murphy et al., 2003). Comparison of these gene lists to the list of intestinal genes identified by our SAGE analysis (McGhee et al., 2007) indicates that, overall, the daf-2/daf-16 responsive genes do not appear to be preferentially expressed in the adult intestine. However, this conclusion does not preclude particular lifespan-related genes acting in the intestine and there have been two recent important examples of such behaviour. The kri-1 gene acts in the intestine to promote the nuclear localization of DAF-16 in response to a signal from the germline (Berman and Kenyon, 2006). The daf-36 gene encodes an oxygenase that is expressed primarily in the intestine and that has been suggested to perform an early step in the synthesis of a steroid-like ligand used for communicating between different tissues, including the intestine (Rottiers et al., 2006).
The C. elegans intestine is one of the few cell lineages where a plausible sequence of direct molecular interactions can be proposed throughout the life cycle, beginning with maternally-derived factors in the cytoplasm of the fertilized zygote, progressing through a small number of zygotic transcription factors and ending with the transcription of vitellogenin genes in the adult intestine to provide food for the next generation. In the current section, we emphasize steps that occur after endoderm specification, i.e., the transcription factors involved with intestinal differentiation, function and maintenance.
Beginning with the early work on vitellogenin genes by Blumenthal and coworkers, promoters of the following genes, expressed exclusively or primarily in the intestine and at a range of developmental stages, have been analyzed to the point where specific cis-acting sites have been mutated: vit-2 (MacMorris and Blumenthal, 1993; MacMorris et al., 1994); cpr-1 (Britton et al., 1998); ges-1 (Egan et al., 1995; Marshall and McGhee, 2001); mtl-1 and mtl-2 (Moilanen et al., 1999); pho-1 (Fukushige et al., 2005); spl-1 (Oskouian et al., 2005); elo-6 (Pauli et al., 2006) and three genes encoding the enzmyes of polyamine synthesis (Luersen et al., 2004). For all of the genes investigated, an extended GATA-like site is critically important for intestine-specific transcription. This conclusion has been confirmed by computational analyses of intestine gene promoters (McGhee et al., 2007; Pauli et al., 2006). Only a small number of non-GATA sites have been experimentally implicated in the control of intestine gene transcription: the VPE1/TGTCAAT elements in vitellogenin promoters (MacMorris et al., 1992; MacMorris et al., 1994), MAB-3 binding sites in vitellogenin promoters (Yi and Zarkower, 1999) and a SKN-1 binding site in the promoter of the gcs-1 gene (An and Blackwell, 2003). In the case of the gcs-1 gene, the critical SKN-1 site overlaps with a GATA site so it remains possible that a GATA factor is also involved here as well. A plausible interaction between MAB-3 and GATA sites in regulating vitellogenin transcription will be discussed below.
The C. elegans genome encodes eleven zinc-finger GATA-related factors, compared to six GATA-factors in vertebrates (Lowry, 2000; Patient, 2002). The crucial involvement of several of these GATA factors in the specification of the endoderm will be discussed in the future WormBook chapter by J. Rothman and M. Maduro and is only briefly summarized here.
Specification of the C. elegans endoderm begins with the maternally-derived bZIP-like SKN-1 factor determining the fate of the EMS blastomere of the four cell embryo (see Figure 2; Bowerman et al., 1992; Bowerman et al., 1993). The EMS blastomere then divides asymmetrically to produce the anterior MS cell (mesoderm) and posterior E cell (endoderm), the clonal progenitor of the intestine. The crucial asymmetry of this division is introduced by a signal passed to the E cell from the adjacent posterior cell P2 (Goldstein, 1992; Goldstein, 1993); this Wnt-MAPK-Src-pathway-based signal (Rocheleau et al., 1997; Thorpe et al., 1997) leads to a lowering of the nuclear concentration of the HMG protein POP-1 (Lin et al., 1995; Lin et al., 1998; Lo et al., 2004), thereby relieving repression of the endoderm pathway and, at the same time converting POP-1 (or a fraction of POP-1) into an endoderm gene activator (Maduro et al., 2005; Shetty et al., 2005). Together with POP-1 in its role as transcriptional activator, SKN-1 also directly activates transcription of genes encoding the two endoderm-specifying GATA factors END-1 and END-3 (Zhu et al., 1997; Zhu et al., 1998; Maduro et al., 2005; Maduro et al., 2005; Shetty et al., 2005). In the EMS blastomere, SKN-1 is also responsible for activating transcription of genes encoding two small redundant highly-diverged GATA factors called MED-1 and MED-2 (Maduro et al., 2001); although important to MS blastomere fate (Maduro et al., 2001; Broitman-Maduro et al., 2005), the MED-1 and MED-2 factors appear much less important in determining E cell fate (Goszczynski and McGhee, 2005; Captan et al., 2006; Maduro et al., 2006). The homeobox protein PAL-1, whose principal maternal role is to specify the fate of the C and D blastomeres (Hunter and Kenyon, 1996), also contributes to end-1/end-3 activation (Maduro et al., 2005) but perhaps only in the experimentally-induced situation where SKN-1 has been removed. Expression of the end-1 and end-3 genes is transient; transcripts can be detected at the 1E cell stage but have largely decayed by the 8E cell stage (Zhu et al., 1997; Baugh et al., 2005); at the moment, there is no evidence either for or against the perdurance of END-1/3 protein. The current model is that END-1 and END-3 set in motion the entire endoderm specification pathway, one of the central features of which is the activation of genes encoding the next round of GATA factors, chiefly ELT-2.
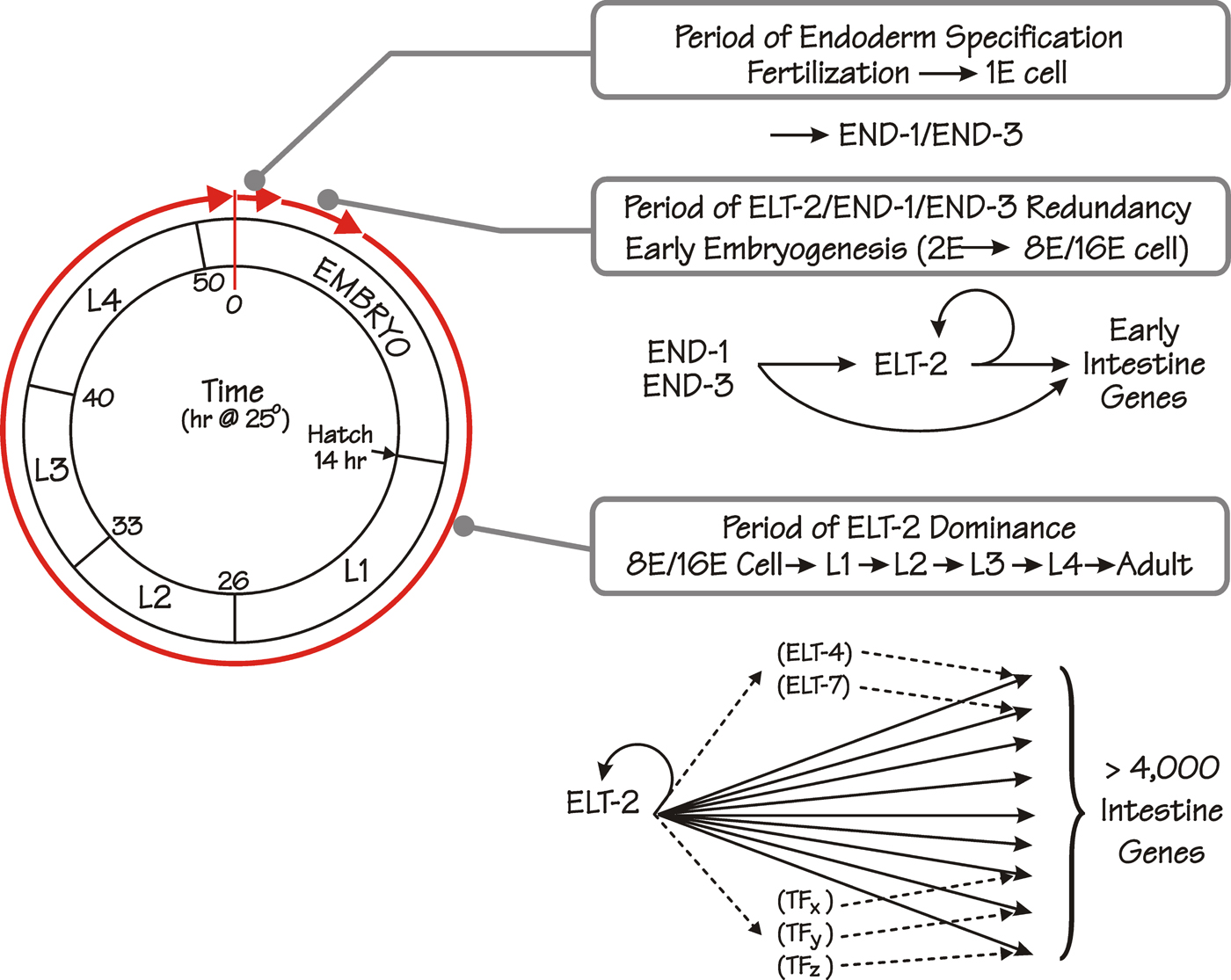
The elt-2 gene (where elt stands for erythrocyte-like transcription factor (sic); Spieth et al., 1991) encodes a single-zinc-finger GATA-type transcription factor (Hawkins and McGhee, 1995) that, we suggest, is the predominant transcription factor in the C. elegans intestine following the early steps of endoderm specification (McGhee et al., 2007); see Figure 5. ELT-2 expression begins during the 2E cell stage and persists into adulthood; maintenance of correct ELT-2 levels likely involves direct elt-2 gene autoregulation (Fukushige et al., 1998; Fukushige et al., 1999). Deletion of elt-2 is lethal: the intestinal lumen is blocked (the gut-obstructed or Gob phenotype; Fukushige et al., 1998) and the afflicted animals die as hatched L1 larvae, presumably from starvation. Nonetheless, the elt-2(-/-) intestine is reasonably well-formed and, at a casual glance, almost normal. Thus, other transcription factors (possibly only END-1 and END-3) must be capable of building most of the early embryonic intestine. However, ELT-2 probably contributes to this early phase of intestine development because it is sufficient (though not necessary) for expression of early endoderm markers such as ges-1, ifb-2 and gut granules (Fukushige et al., 1998). The essential functions of ELT-2 are first revealed later in embryogenesis. In particular, ELT-2 has been shown to be necessary for the expression of three intestine-specific genes first expressed late in embryogenesis and analyzed immediately following hatching: the mtl-2 metallothionein gene (Moilanen et al., 1999), the spl-1 sphingosine phosphate lyase gene (Oskouian et al., 2005) and the pho-1 intestinal acid phosphatase gene (Fukushige et al., 2005).
|
Figure 5. The proposed role(s) of the ELT-2 GATA factor in the overall pathway forming the C. elegans endoderm. The successive life stages and the approximate times (hours at 25°C following fertilization) of the C. elegans life cycle are depicted by the circle on the left (adapted from (Wood et al., 1980). The first stage in endoderm formation occupies ~1.5 hours following fertilization, ending with production of END-1 and END-3 in the E blastomere, the defining event in endoderm specification. The second phase (“Period of ELT-2/END-1/END-3 Redundancy”) begins at the 2E cell stage when ELT-2 is first produced and ends at the 8E-16E cell stage (~4 hours after fertilization) when END-1 and END-3 levels have decayed. We suggest that ELT-2, END-1 and END-3 all participate in the transcriptional activation of intestine genes during this early phase of intestine development. The third phase (“Period of ELT-2 Dominance”) begins at the 8E-16E cell stage and continues throughout all subsequent larval stages including the adult. In this phase, we propose that ELT-2 is directly and necessarily involved in all acts of transcription in the intestine, including transcription of genes encoding other transcription factors (e.g. ELT-4, ELT-7, TFx, TFy etc), which in turn may cooperate with ELT-2 in mounting particular transcriptional responses. ELT-7 (and possibly ELT-4) may provide redundant backup for a minor fraction of genes regulated by ELT-2, i.e. an elt-7; elt-2 double knockout has a slightly more severe phenotype than does an elt-2 knockout by itself (unpublished results of K. Strohmaier and J. Rothman; our unpublished results). Figure and figure legend reproduced from McGhee et al. (2006) with permission from Elsevier.
ELT-4 and ELT-7 (C18G1.2) are two other GATA factors expressed in the developing intestine beginning approximately mid-embryogenesis (Fukushige et al., 2003) and unpublished results of K. Strohmaier cited in (Maduro and Rothman, 2002). However, neither factor seems to have a major influence on intestine development, i.e., animals in which the elt-4 gene (Fukushige et al., 2003) or the elt-7 gene (Fukushige et al., 2005; Oskouian et al., 2005 and unpublished results of Keith Strohmaier) or both elt-4 and elt-7 genes (McGhee et al., 2007) have either been deleted or their transcripts ablated by RNAi are essentially wildtype. Nonetheless, elt-2 elt-7 double knockouts show a slightly more severe phenotype than does the elt-2 single knockout (unpublished results of Keith Strohmaier; our unpublished results), suggesting that elt-7 is partially redundant with elt-2.
We thus propose the simple model (see Figure 5) that, following endoderm specification, ELT-2 participates in all acts of transcription in the intestine. However, other transcription factors are clearly present in the intestine (see below) and are likely to modulate the action of ELT-2 in different developmental or environmental circumstances. Transcription of the vitellogenin genes in the hermaphrodite intestine provides a possible example how this could occur. The vitellogenin (yolk protein) genes are expressed in the adult hermaphrodite intestine but not in the male intestine (Kimble and Sharrock, 1983). It had been known for some time that the mab-3 gene is involved in repressing male vitellogenin synthesis (Shen and Hodgkin, 1988) and Yi and Zarkower have shown that this effect is direct (Yi and Zarkower, 1999). The MAB-3 protein shows similarity to Drosophila doublesex, which, among other things, is also responsible for sex-specific regulation of yolk protein synthesis in flies (Raymond et al., 1998). The preferred MAB-3 binding sequence was identified in vitro and related sequences were then identified in the promoters of the half-dozen vitellogenin genes (Yi and Zarkower, 1999). MAB-3 binds directly to one such site in an abbreviated promoter-reporter construct from the vit-2 gene and mutation of this site causes reporter gene derepression in male intestines. A critical GATA site (MacMorris et al., 1992; MacMorris et al., 1994) lies immediately adjacent to this MAB-3 site, suggesting that MAB-3 could act in the male intestine by repressing the activity of ELT-2. GATA-like sites also lie in the vicinity of MAB-3 sites found in other vitellogenin gene promoters (MacMorris et al., 1992; MacMorris et al., 1994; Yi and Zarkower, 1999).
Analysis of a SAGE library prepared from FACS-sorted embryonic intestine cells identifies dozens of different transcription factors (D. G. Moerman and JDM, in preparation) but only a small number have been investigated experimentally.
tbx-8 and tbx-9 are highly similar T-box genes expressed in the embryo posterior (intestine, muscles and hypodermis). Expression in the intestine can be detected in the 2E and 4E cells but declines thereafter. Most phenotypes of tbx-8 and tbx-9 double knockouts are associated with hypodermis and muscle but cells in the mutant intestine appear mis-positioned and clumped (Pocock et al., 2004).
The forkhead factor PHA-4 is critical for embryonic formation of the pharynx and the rectum (Mango et al., 1994; Azzaria et al., 1996; Horner et al., 1998; Kalb et al., 1998). However, pha-4 is also expressed in the intestine at all stages; rather subtle morphological changes can be detected in the embryonic intestine of pha-4 mutants. Intestinal expression of pha-4 may well be controlled by ELT-2 (Kalb et al., 1998).
The UNC-130 protein is a forkhead transcription factor best known for its role in regulating cell migration (Nash et al., 2000). unc-130 is expressed in a variety of tissues, among which is the intestinal primordium. Intestinal expression begins at the approximately 8E cell stage and persists into adulthood (Nash et al., 2000).
The ceh-13 homeobox gene is expressed in the posterior cells of the intestinal primordium (Wittmann et al., 1997), among several other places in the embryo. It is not clear what causes this patterned expression nor whether ceh-13 expression has any consequences for overall anterior-patterning in the intestine. As mentioned earlier, the pal-1 homeobox gene is expressed in the int5 cells of the developing intestine primordium (Edgar et al., 2001).
The die-1 gene encodes a zinc-finger protein, possibly a transcription factor, that was identified because mutants show aberrant morphogenesis of the hypodermis (Heid et al., 2001). However, die-1 is expressed widely in the embryo, including in the developing intestine; die-1 mutants show intestinal defects in which the intestine does not attach to the pharynx or to the rectum.
NHR-25, the C. elegans homolog of FtzF1, has an early phase of expression restricted to the E lineage (approximately 4E cell stage to comma stage) but subsequently becomes expressed more widely (Asahina et al., 2000). Knockout of NHR-25 causes embryonic arrest but the intestine primordium still expresses differentiation markers. HNF-4 type nuclear hormone receptors (like NHR-25) are usually associated with endoderm in other animals but this does not appear to be true of the same family of factors in C. elegans (Robinson-Rechavi et al., 2005).
SAGE analysis of adult intestines identifies transcripts corresponding to over a hundred different transcription factors (McGhee et al., 2007) but, like the transcription factors in the embryonic intestine, only a small number have been investigated experimentally. However, one important conclusion is that the majority of these transcription factors do not show an RNAi phenotype, at least in high throughput screens. Of the approximately 10% of adult intestine transcription factors that do show an RNAi phenotype, none except ELT-2 show a major developmental phenotype centred on the intestine.
Control of lipid metabolism and lipid homeostasis is a burgeoning field of study in C. elegans, holding promise for understanding lipid-related diseases in humans. Several transcription factors controlling specific genes in, for example, fatty acid metabolism, have been identified and the emerging view is of complex redundant overlapping transcriptional control, centred on the intestine but acting in other tissues as well. Three points of interest are:
The nuclear hormone receptor NHR-49 (a candidate PPAR homologue) is expressed in a number of different tissues including the intestine. Knockout of NHR-49 leads to enhanced fat storage in the intestine (and presumably other tissues as well). A number of potential targets have been identified, in particular genes involved in mitochondrial β-oxidation of fatty acids and genes involved in fatty acid desaturation (Gilst et al., 2005).
The nhr-80 gene has also been shown to be involved in controlling genes involved in fatty acid desaturation, in particular the fat-5, fat-6 and fat-7 genes encoding Δ9-desaturases (Brock et al., 2006). fat-7 is expressed exclusively in the intestine and expression is abolished in nhr-80 mutants. fat-5 and fat-6 are expressed strongly but not exclusively in the intestine; their expression levels are decreased (but not abolished) by loss of nhr-80. As noted earlier, simultaneous loss of all three fat-5, fat-6 and fat-7 genes is lethal but when only fat-6 or fat-7 are lost, the remaining genes are able to adjust their levels of expression. This ability to compensate by adjusting expression levels is controlled by nhr-80 (Brock et al., 2006).
LPD-1 is the C. elegans homolog of mammalian SREBP (sterol response element binding protein) and is expressed exclusively in the intestine. A number of potential direct downstream targets have been identified and one such target (lpd-3) has also been shown to be exclusively expressed in the intestine (McKay et al., 2003). The elo-5 and elo-6 genes encode fatty acid elongation enzymes, and are good candidates to be direct targets of LPD-1 (Kniazeva et al., 2004). Intestinal expression of elo-6 is largely abolished by mutation of a GATA site in the promoter (Pauli et al., 2006) suggesting that regulation of elo-6 might involve cooperation between ELT-2 and LPD-1.
We note the following additional points of interest concerning intestinal transcription factors:
Overall, nuclear hormone receptors are highly represented among transcription factors expressed at the highest levels in the adult intestine. In addition to the nhr-49 and nhr-80 genes described above, nhr-8 is expressed exclusively in the intestine and loss of nhr-8 function enhances sensitivity to toxins such as colchicine (Lindblom et al., 2001).
The UNC-62 homeobox factor is the C. elegans homolog of Drosophila homothorax and unc-62 transcripts are present at high levels in the adult intestine (McGhee et al., 2007). unc-62 is expressed widely in the embryo and unc-62 mutants are embryonic lethal (Van Auken et al., 2002).
CEH-20 is the C. elegans homolog of Drosophila extradenticles (Van Auken et al., 2002) and transcripts can be detected at moderate levels in the adult intestine (McGhee et al., 2007).
Transcripts corresponding to pha-4, the forkhead “organ identity factor” for the pharynx, can also be detected in the adult intestine (McGhee et al., 2007).
The forkhead factor DAF-16 is expressed widely throughout the worm and its potential role in the intestine was described above in the section on aging.
In the preceding section, we have focused on transcription factors that bind to specific DNA sequences in the regulatory regions of intestinal genes. However, there are dozens and dozens of other factors that are usually referred to as General Transcription Factors, either involved with the basic transcriptional machinery or in providing links between this basic machinery and the promoter specific factors. This important subject is covered in the WormBook chapter by Blackwell and Walker (see Transcription mechanisms). Of particular interest to understanding control of gene expression in the C. elegans intestine are two recent studies that show that two transcription factors involved in the control of lipid metabolism, nhr-49 and sbp-1/lpd-1 interact with a particular subunit of the mediator complex; the mediator subunit involved in controlling these genes in C. elegans (MDT-15) is the homolog of the mediator subunit controlling the homologous genes in humans (Taubert et al., 2006; Yang et al., 2006)
Finally, there are increasing hints how transcription in the intestine might also be subject to “higher” control, possibly including regulation of chromatin structure. For example, members of the C. elegans Polycomb-like group of factors, in particular MES-2 and MES-6, are prominently expressed in the intestine (Ross and Zarkower, 2003). Also, screens for SynMuv (synthetic multivulva) genes, some of which have been found to be involved in higher order control of gene regulation, can be detected because of their permissive effects on intestine gene expression (Poulin et al., 2005). Suppressors of SynMuv genes have also been identified, many of which reverse SynMuv effects on ectopic expression of reporter genes in the intestine (Cui et al., 2006). It is not yet known how these genes/factors/mechanisms are involved in normal processes of gene expression in the intestine but they point to complex additional layers of regulation.
It is clear that the above descriptions have only touched the surface of intestinal biology in C. elegans but they do lead to the following set of (rather obvious) questions that must now be addressed:
What fraction of the food-derived macromolecules is taken up by endocytosis and what fraction is digested within the intestinal lumen, with degradation products assimilated via dedicated transporters? How can C. elegans ingest and digest so many bacteria so quickly?
At the molecular level, how does such a spectacular sub-cellular structure as the brush border assemble, with all microvilli the same length?
How does the intestine communicate with the outside environment or with other tissues such as the gonad? There have been several important recent insights into such communication pathways (Berman and Kenyon, 2006; Mak et al., 2006) but there are certain to be others.
How does the intestine participate in the overall energy economy of the animal and in fundamental biological processes such as regulation of growth and size? Is food distribution from the intestine to the rest of the worm constitutive or “on demand”?
Even if the main transcription factors regulating intestine development have been identified, how do they all work together? Will understanding transcription factors really tell us how C. elegans makes its intestine?
Work in the author's laboratory was funded by the Canadian Institutes of Health Research, the Alberta Heritage Foundation for Medical Research and the Canada Research Chairs. I welcome comments about errors, omissions, mis-interpretations and suggestions for future revisions.
Albert, P.S., and Riddle, D.L. (1988). Mutants of Caenorhabditis elegans that form dauer-like larvae. Dev. Biol. 126, 270–293. Abstract Article
An, J.H., and Blackwell, T.K. (2003). SKN-1 links C. elegans mesendodermal specification to a conserved oxidative stress response. Genes Dev. 17, 1882–1893. Abstract Article
An, J.H., Vranas, K., Lucke, M., Inoue, H., Hisamoto, N., Matsumoto, K., and Blackwell, T.K. (2005). Regulation of the Caenorhabditis elegans oxidative stress defense protein SKN-1 by glycogen synthase kinase-3. Proc. Natl. Acad. Sci. U.S.A. 102, 16275–16280. Abstract Article
Appleford, P.J., Griffiths, M., Yao, S.Y., Ng, A.M., Chomey, E.G., Isaac, R.E., Coates, D., Hope, I.A., Cass, C.E., Young, J.D., and Baldwin, S.A. (2004). Functional redundancy of two nucleoside transporters of the ENT family (CeENT1, CeENT2) required for development of Caenorhabditis elegans. Mol. Membr. Biol. 21, 247–259. Abstract Article
Asahina, M., Ishihara, T., Jindra, M., Kohara, Y., Katsura, I., and Hirose, S. (2000). The conserved nuclear receptor Ftz-F1 is required for embryogenesis, moulting and reproduction in Caenorhabditis elegans. Genes Cells 5, 711–723. Abstract Article
Ashrafi, K., Chang, F.Y., Watts, J.L., Fraser, A.G., Kamath, R.S., Ahringer, J., and Ruvkun, G. (2003). Genome-wide RNAi analysis of Caenorhabditis elegans fat regulatory genes. Nature 421, 268–272. Abstract Article
Avery, L., and Shtonda, B.B. (2003). Food transport in the C. elegans pharynx. J. Exp. Biol. 206, 2441–2457. Abstract Article
Avery, L., and Thomas, J.H. (1997). Feeding and Defecation, In C. elegans II, D.L. Riddle, T. Blumenthal, B.J. Meyer, and J.R. Priess, eds. (Cold Spring Harbor Laboratory Press), pp. 679–716.
Azzaria, M., Goszczynski, B., Chung, M.A., Kalb, J.M., and McGhee, J.D. (1996). A fork head/HNF-3 homolog expressed in the pharynx and intestine of the Caenorhabditis elegans embryo. Dev. Biol. 178, 289–303. Abstract Article
Banyai, L., and Patthy, L. (1998). Amoebapore homologs of Caenorhabditis elegans. Biochim. Biophys. Acta 1429, 259–264. Abstract
Baugh, L.R., Hill, A.A., Claggett, J.M., Hill-Harfe, K., Wen, J.C., Slonim, D.K., Brown, E.L., and Hunter, C.P. (2005). The homeodomain protein PAL-1 specifies a lineage-specific regulatory network in the C. elegans embryo. Development 132, 1843–1854. Abstract Article
Beh, C.T., Ferrari, D.C., Chung, M.A., and McGhee, J.D. (1991). An acid phosphatase as a biochemical marker for intestinal development in the nematode Caenorhabditis elegans. Dev. Biol. 147, 133–143. Abstract Article
Behm, C.A. (2002). Metabolism, In The Biology of Nematodes, D.L. Lee, ed. (London: Taylor&Francis), pp. 261–290.
Berman, J.R., and Kenyon, C. (2006). Germ-cell loss extends C. elegans life span through regulation of DAF-16 by kri-1 and lipophilic-hormone signaling. Cell 124, 1055–1068. Abstract Article
Berman, K., McKay, J., Avery, L., and Cobb, M. (2001). Isolation and characterization of pmk-(1-3): three p38 homologs in Caenorhabditis elegans. Mol. Cell Biol. Res. Commun. 4, 337–344. Abstract Article
Borgonie, G., van Driessche, E., Link, C.D., Claeys, M., de Waele, D., and Coomans, A. (1997). Internal lectin binding patterns in the nematodes Caenorhabditis elegans, Panagrolaimus superbus and Acrobeloides maximus. Fundam. Appl. Nematol. 20, 173–186.
Borgonie, G., Claeys, M., Leyns, F., Arnaut, G., de Waele, D., and Coomans, A. (1996). Effect of nematicidal Bacillus thuringiensis strains on free-living nematodes. Fundam. Appl. Nematol. 19, 391–398.
Borgonie, G., Claeys, M., Vanfleteren, J., de Waele, D., and Coomans, A. (1995). Presence of peritrophic-like membranes in the intestine of three bacteriophagous nematdoes (Nematoda: Rhabdita). Fundam. Appl. Nematol. 18, 227–233. Abstract Article
Borgonie, G., Van Driessche, R., Leyns, F., Arnaut, G., De Waele, D., and Coomans, A. (1995). Germination of Bacillus thuringiensis spores in bacteriophagous nematodes (Nematoda: Rhabditida). J. Invertebr. Pathol. 65, 61–67. Abstract Article
Borgonie, G., van Driessche, E., Link, C.D., de Waele, D., and Coomans, A. (1994). Tissue treatment for whole mount internal lectin staining in the nematodes Caenorhabditis elegans, Panagrolaimus superbus and Acrobeloides maximus. Histochemistry 101, 379–384. Abstract Article
Bossinger, O., Fukushige, T., Claeys, M., Borgonie, G., and McGhee, J.D. (2004). The apical disposition of the Caenorhabditis elegans intestinal terminal web is maintained by LET-413. Dev. Biol. 268, 448–456. Abstract Article
Bossinger, O., Klebes, A., Segbert, C., Theres, C., and Knust, E. (2001). Zonula adherens formation in Caenorhabditis elegans requires dlg-1, the homologue of the Drosophila gene discs large. Dev. Biol. 230, 29–42. Abstract Article
Bossinger, O., and Schierenberg, E. (1996). The use of fluorescent marker dyes for studying intercellular communication in nematode embryos. Int. J. Dev. Biol. 40, 431–439. Abstract
Bowerman, B., Draper, B.W., Mello, C.C., and Priess, J.R. (1993). The maternal gene skn-1 encodes a protein that is distributed unequally in early C. elegans embryos. Cell 74, 443–452. Abstract Article
Bowerman, B., Eaton, B.A., and Priess, J.R. (1992). skn-1, a maternally expressed gene required to specify the fate of ventral blastomeres in the early C. elegans embryo. Cell 68, 1061–1075. Abstract Article
Boxem, M., and van den Heuvel, S. (2001). lin-35 Rb and cki-1 Cip/Kip cooperate in developmental regulation of G1 progression in C. elegans. Development 128, 4349–4359. Abstract
Braeckman, B.P., Demetrius, L., and Vanfleteren, J.R. (2006). The dietary restriction effect in C. elegans and humans: is the worm a one-millimeter human? Biogerontology. Abstract Article
Branicky, R., and Hekimi, S. (2006). What keeps C. elegans regular: the genetics of defecation. Trends Genet. 22, 571–579. Abstract Article
Britton, C., McKerrow, J.H., and Johnstone, I.L. (1998). Regulation of the Caenorhabditis elegans gut cysteine protease gene cpr-1: requirement for GATA motifs. J. Mol. Biol. 283, 15–27. Abstract Article
Britton, C., and Murray, L. (2006). Using Caenorhabditis elegans for functional analysis of genes of parasitic nematodes. Int. J. Parasitol. 36, 651–659. Abstract Article
Brock, T.J., Browse, J., and Watts, J.L. (2006). Genetic regulation of unsaturated fatty acid composition in C. elegans. PLoS Genet. 2, e108. Abstract Article
Broeks, A., Gerrard, B., Allikmets, R., Dean, M., and Plasterk, R.H. (1996). Homologues of the human multidrug resistance genes MRP and MDR contribute to heavy metal resistance in the soil nematode Caenorhabditis elegans. EMBO J. 15, 6132–6143. Abstract
Broeks, A., Janssen, H.W., Calafat, J., and Plasterk, R.H. (1995). A P-glycoprotein protects Caenorhabditis elegans against natural toxins. EMBO J. 14, 1858–1866. Abstract
Broitman-Maduro, G., Maduro, M.F., and Rothman, J.H. (2005). The noncanonical binding site of the MED-1 GATA factor defines differentially regulated target genes in the C. elegans mesendoderm. Dev. Cell 8, 427–433. Abstract Article
Brooks, D.R., Hooper, N.M., and Isaac, R.E. (2003). The Caenorhabditis elegans orthologue of mammalian puromycin-sensitive aminopeptidase has roles in embryogenesis and reproduction. J. Biol. Chem. 278, 42795–42801. Abstract Article
Captan, V.V., Goszczynski, B., and McGhee, J.D. (2006). Neither maternal nor zygotic med-1/med-2 genes play a major role in specifying the C. elegans endoderm. Genetics (in press).
Chen, C.C., Schweinsberg, P.J., Vashist, S., Mareiniss, D.P., Lambie, E.J., and Grant, B.D. (2006). RAB-10 is required for endocytic recycling in the Caenorhabditis elegans intestine. Mol. Biol. Cell 17, 1286–1297. Abstract Article
Chen, S., Zhou, S., Sarkar, M., Spence, A.M., and Schachter, H. (1999). Expression of three Caenorhabditis elegans N-acetylglucosaminyltransferase I genes during development. J. Biol. Chem. 274, 288–297. Abstract Article
Choy, R.K., Kemner, J.M., and Thomas, J.H. (2006). Fluoxetine-resistance genes in Caenorhabditis elegans function in the intestine and may act in drug transport. Genetics 172, 885–892. Abstract Article
Choy, R.K., and Thomas, J.H. (1999). Fluoxetine-resistant mutants in C. elegans define a novel family of transmembrane proteins. Mol. Cell 4, 143–152. Abstract Article
Church, D.L., and Lambie, E.J. (2003). The promotion of gonadal cell divisions by the Caenorhabditis elegans TRPM cation channel GON-2 is antagonized by GEM-4 copine. Genetics 165, 563–574. Abstract
Clokey, G.V., and Jacobson, L.A. (1986). The autofluorescent "lipofuscin granules" in the intestinal cells of Caenorhabditis elegans are secondary lysosomes. Mech. Ageing Dev. 35, 79–94. Abstract Article
Clucas, C., Cabello, J., Bussing, I., Schnabel, R., and Johnstone, I.L. (2002). Oncogenic potential of a C. elegans cdc25 gene is demonstrated by a gain-of-function allele. EMBO J. 21, 665–674. Abstract Article
Costa, M., Raich, W., Agbunag, C., Leung, B., Hardin, J., and Priess, J.R. (1998). A putative catenin-cadherin system mediates morphogenesis of the Caenorhabditis elegans embryo. J. Cell. Biol. 141, 297–308. Abstract Article
Croce, A., Cassata, G., Disanza, A., Gagliani, M.C., Tacchetti, C., Malabarba, M.G., Carlier, M.F., Scita, G., Baumeister, R., and Di Fiore, P.P. (2004). A novel actin barbed-end-capping activity in EPS-8 regulates apical morphogenesis in intestinal cells of Caenorhabditis elegans. Nat. Cell Biol. 6, 1173–1179. Abstract Article
Cui, M., Kim, E.B., and Han, M. (2006). Diverse chromatin remodeling genes antagonize the Rb-involved SynMuv pathways in C. elegans. PLoS Genet. 2, e74. Abstract Article
Dal Santo, P., Logan, M.A., Chisholm, A.D., and Jorgensen, E.M. (1999). The inositol trisphosphate receptor regulates a 50-second behavioral rhythm in C. elegans. Cell 98, 757–767. Abstract Article
de Voer, G., van der Bent, P., Rodrigues, A.J., van Ommen, G.J., Peters, D.J., and Taschner, P.E. (2005). Deletion of the Caenorhabditis elegans homologues of the CLN3 gene, involved in human juvenile neuronal ceroid lipofuscinosis, causes a mild progeric phenotype. J. Inherit. Metab. Dis. 28, 1065–1080. Abstract Article
Deppe, U., Schierenberg, E., Cole, T., Krieg, C., Schmitt, D., Yoder, B., and von Ehrenstein, G. (1978). Cell lineages of the embryo of the nematode Caenorhabditis elegans. Proc. Natl. Acad. Sci. U.S.A. 75, 376–380. Abstract Article
Ding, L., and Candido, E.P. (2000). Association of several small heat-shock proteins with reproductive tissues in the nematode Caenorhabditis elegans. Biochem. J. 351, 13–17. Abstract Article
Dodemont, H., Riemer, D., Ledger, N., and Weber, K. (1994). Eight genes and alternative RNA processing pathways generate an unexpectedly large diversity of cytoplasmic intermediate filament proteins in the nematode Caenorhabditis elegans. EMBO J. 13, 2625–2638. Abstract
Dodemont, H., Riemer, D., and Weber, K. (1990). Structure of an invertebrate gene encoding cytoplasmic intermediate filament (IF) proteins: implications for the origin and the diversification of IF proteins. EMBO J. 9, 4083–4094. Abstract
Dong, J., Song, M.O., and Freedman, J.H. (2005). Identification and characterization of a family of Caenorhabditis elegans genes that is homologous to the cadmium-responsive gene cdr-1. Biochim. Biophys. Acta 1727, 16–26. Abstract Article
Edgar, L.G., Carr, S., Wang, H., and Wood, W.B. (2001). Zygotic expression of the caudal homolog pal-1 is required for posterior patterning in Caenorhabditis elegans embryogenesis. Dev. Biol. 229, 71–88. Abstract Article
Edgar, L.G., and McGhee, J.D. (1988). DNA synthesis and the control of embryonic gene expression in C. elegans. Cell 53, 589–599. Abstract Article
Egan, C.R., Chung, M.A., Allen, F.L., Heschl, M.F., Van Buskirk, C.L., and McGhee, J.D. (1995). A gut-to-pharynx/tail switch in embryonic expression of the Caenorhabditis elegans ges-1 gene centers on two GATA sequences. Dev. Biol. 170, 397–419. Abstract Article
Espelt, M.V., Estevez, A.Y., Yin, X., and Strange, K. (2005). Oscillatory Ca2+ signaling in the isolated Caenorhabditis elegans intestine: role of the inositol-1,4,5-trisphosphate receptor and phospholipases C beta and gamma. J. Gen. Physiol. 126, 379–392. Abstract Article
Estevez, A.Y., Roberts, R.K., and Strange, K. (2003). Identification of store-independent and store-operated Ca2+ conductances in Caenorhabditis elegans intestinal epithelial cells. J. Gen. Physiol. 122, 207–223. Abstract Article
Estevez, A.Y., and Strange, K. (2005). Calcium feedback mechanisms regulate oscillatory activity of a TRP-like Ca2+ conductance in C. elegans intestinal cells. J. Physiol. 567, 239–251. Abstract Article
Fares, H., and Grant, B. (2002). Deciphering endocytosis in Caenorhabditis elegans. Traffic 3, 11–19. Abstract Article
Fei, Y.J., Inoue, K., and Ganapathy, V. (2003). Structural and functional characteristics of two sodium-coupled dicarboxylate transporters (ceNaDC1 and ceNaDC2) from Caenorhabditis elegans and their relevance to life span. J. Biol. Chem. 278, 6136–6144. Abstract Article
Fei, Y.J., Liu, J.C., Inoue, K., Zhuang, L., Miyake, K., Miyauchi, S., and Ganapathy, V. (2004). Relevance of NAC-2, an Na+-coupled citrate transporter, to life span, body size and fat content in Caenorhabditis elegans. Biochem. J. 379, 191–198. Abstract Article
Feinberg, E.H., and Hunter, C.P. (2003). Transport of dsRNA into cells by the transmembrane protein SID-1. Science 301, 1545–1547. Abstract Article
Francis, G.R., and Waterston, R.H. (1985). Muscle organization in Caenorhabditis elegans: localization of proteins implicated in thin filament attachment and I-band organization. J. Cell Biol. 101, 1532–1549. Abstract Article
Freedman, J.H., Slice, L.W., Dixon, D., Fire, A., and Rubin, C.S. (1993). The novel metallothionein genes of Caenorhabditis elegans. Structural organization and inducible, cell-specific expression. J. Biol. Chem. 268, 2554–2564. Abstract
Fukushige, T., Goszczynski, B., Tian, H., and McGhee, J.D. (2003). The evolutionary duplication and probable demise of an endodermal GATA factor in Caenorhabditis elegans. Genetics 165, 575–588. Abstract
Fukushige, T., Goszczynski, B., Yan, J., and McGhee, J.D. (2005). Transcriptional control and patterning of the pho-1 gene, an essential acid phosphatase expressed in the C. elegans intestine. Dev. Biol. 279, 446–461. Abstract Article
Fukushige, T., Hawkins, M.G., and McGhee, J.D. (1998). The GATA-factor elt-2 is essential for formation of the Caenorhabditis elegans intestine. Dev. Biol. 198, 286–302. Abstract
Fukushige, T., Hendzel, M.J., Bazett-Jones, D.P., and McGhee, J.D. (1999). Direct visualization of the elt-2 gut-specific GATA factor binding to a target promoter inside the living Caenorhabditis elegans embryo. Proc. Natl. Acad. Sci. U.S.A. 96, 11883–11888. Abstract Article
Fukuyama, M., Gendreau, S.B., Derry, W.B., and Rothman, J.H. (2003). Essential embryonic roles of the CKI-1 cyclin-dependent kinase inhibitor in cell-cycle exit and morphogenesis in C. elegans. Dev. Biol. 260, 273–286. Abstract Article
Geier, G., Banaj, H.J., Heid, H., Bini, L., Pallini, V., and Zwilling, R. (1999). Aspartyl proteases in Caenorhabditis elegans. Isolation, identification and characterization by a combined use of affinity chromatography, two-dimensional gel electrophoresis, microsequencing and databank analysis. Eur. J. Biochem. 264, 872–879. Abstract Article
Ghafouri, S., and McGhee, J.D. (2007). Bacterial residence time in the intestine of Caenorhabditis elegans Nematology (in press).
Ghenea, S., Takeuchi, M., Motoyama, J., Sasamoto, K., Kunau, W.-H., Kamiryo, T., and Bun-Ya, M. (2001). The cDNA Sequence and Expression of the AAA-family Peroxin Genes pex-1 and pex-6 from the Nematode Caenorhabditis elegans. Zool. Sci. 18, 675–681. Article
Gilst, M.R., Hadjivassiliou, H., Jolly, A., and Yamamoto, K.R. (2005). Nuclear Hormone Receptor NHR-49 Controls Fat Consumption and Fatty Acid Composition in C. elegans. PLoS Biol 3, e53. Abstract Article
Gobel, V., Barrett, P.L., Hall, D.H., and Fleming, J.T. (2004). Lumen morphogenesis in C. elegans requires the membrane-cytoskeleton linker erm-1. Dev. Cell 6, 865–873. Abstract Article
Goldstein, B. (1992). Induction of gut in Caenorhabditis elegans embryos. Nature 357, 255–257. Abstract Article
Goldstein, B. (1993). Establishment of gut fate in the E lineage of C. elegans: the roles of lineage-dependent mechanisms and cell interactions. Development 118, 1267–1277. Abstract
Goszczynski, B., and McGhee, J.D. (2005). Reevaluation of the Role of the med-1 and med-2 Genes in Specifying the Caenorhabditis elegans Endoderm. Genetics 171, 545–555. Abstract Article
Graham, P.L., Johnson, J.J., Wang, S., Sibley, M.H., Gupta, M.C., and Kramer, J.M. (1997). Type IV collagen is detectable in most, but not all, basement membranes of Caenorhabditis elegans and assembles on tissues that do not express it. J. Cell. Biol. 137, 1171–1183. Abstract Article
Grant, B., and Greenwald, I. (1997). Structure, function, and expression of SEL-1, a negative regulator of LIN-12 and GLP-1 in C. elegans. Development 124, 637–644. Abstract
Griffitts, J.S., Haslam, S.M., Yang, T., Garczynski, S.F., Mulloy, B., Morris, H., Cremer, P.S., Dell, A., Adang, M.J., and Aroian, R.V. (2005). Glycolipids as receptors for Bacillus thuringiensis crystal toxin. Science 307, 922–925. Abstract Article
Griffitts, J.S., Huffman, D.L., Whitacre, J.L., Barrows, B.D., Marroquin, L.D., Muller, R., Brown, J.R., Hennet, T., Esko, J.D., and Aroian, R.V. (2003). Resistance to a bacterial toxin is mediated by removal of a conserved glycosylation pathway required for toxin-host interactions. J. Biol. Chem. 278, 45594–45602. Abstract Article
Griffitts, J.S., Whitacre, J.L., Stevens, D.E., and Aroian, R.V. (2001). Bt toxin resistance from loss of a putative carbohydrate-modifying enzyme. Science 293, 860–864. Abstract Article
Gurvitz, A., Langer, S., Piskacek, M., Hamilton, B., Ruis, H., and Hartig, A. (2000). Predicting the function and subcellular location of Caenorhabditis elegans proteins similar to Saccharomyces cerevisiae beta-oxidation enzymes. Yeast 17, 188–200. Abstract
Hagen, F.K., Layden, M., Nehrke, K., Gentile, K., Berbach, K., Tsao, C.C., and Forsythe, M. (2001). Mucin-Type O-Glycosylation in C. elegans is Initiated by a Family of Glycosyl transferases. Trends Glycosci. Glycotechnol. 13, 463–479.
Hanover, J.A., Forsythe, M.E., Hennessey, P.T., Brodigan, T.M., Love, D.C., Ashwell, G., and Krause, M. (2005). A Caenorhabditis elegans model of insulin resistance: altered macronutrient storage and dauer formation in an OGT-1 knockout. Proc. Natl. Acad. Sci. U.S.A. 102, 11266–11271. Abstract Article
Hawkins, M.G., and McGhee, J.D. (1995). elt-2, a second Gata factor from the nematode Caenorhabditis elegans. J. Biol. Chem. 270, 14666–14671. Abstract Article
Hedgecock, E.M., and White, J.G. (1985). Polyploid tissues in the nematode Caenorhabditis elegans. Dev. Biol. 107, 128–133. Abstract Article
Heid, P.J., Raich, W.B., Smith, R., Mohler, W.A., Simokat, K., Gendreau, S.B., Rothman, J.H., and Hardin, J. (2001). The zinc finger protein DIE-1 is required for late events during epithelial cell rearrangement in C. elegans. Dev. Biol. 236, 165–180 Abstract Article
Hermann, G.J., Leung, B., and Priess, J.R. (2000). Left-right asymmetry in C. elegans intestine organogenesis involves a LIN-12/Notch signaling pathway. Development 127, 3429–3440. Abstract
Hermann, G.J., Schroeder, L.K., Hieb, C.A., Kershner, A.M., Rabbitts, B.M., Fonarev, P., Grant, B.D., and Priess, J.R. (2005). Genetic analysis of lysosomal trafficking in Caenorhabditis elegans. Mol. Biol. Cell 16, 3273–3288. Abstract Article
Hevelone, J., and Hartman, P.S. (1988). An endonuclease from Caenorhabditis elegans: partial purification and characterization. Biochem. Genet. 26, 447–461. Abstract Article
Hirsh, D., Oppenheim, D., and Klass, M. (1976). Development of the reproductive system of Caenorhabditis elegans. Dev. Biol. 49, 200–219. Abstract Article
Hong, Y., Lee, R.C., and Ambros, V. (2000). Structure and function analysis of LIN-14, a temporal regulator of postembryonic developmental events in Caenorhabditis elegans. Mol. Cell Biol. 20, 2285–2295. Abstract Article
Hong, Y., Roy, R., and Ambros, V. (1998). Developmental regulation of a cyclin-dependent kinase inhibitor controls postembryonic cell cycle progression in Caenorhabditis elegans. Development 125, 3585–3597. Abstract
Horner, M.A., Quintin, S., Domeier, M.E., Kimble, J., Labouesse, M., and Mango, S.E. (1998). pha-4, an HNF-3 homolog, specifies pharyngeal organ identity in Caenorhabditis elegans. Genes Dev. 12, 1947–1952. Abstract
Hsu, A.L., Murphy, C.T., and Kenyon, C. (2003). Regulation of aging and age-related disease by DAF-16 and heat-shock factor. Science 300, 1142–1145. Abstract Article
Huang, C.C., Hall, D.H., Hedgecock, E.M., Kao, G., Karantza, V., Vogel, B.E., Hutter, H., Chisholm, A.D., Yurchenco, P.D., and Wadsworth, W.G. (2003). Laminin alpha subunits and their role in C. elegans development. Development 130, 3343–3358. Abstract Article
Huffman, D.L., Abrami, L., Sasik, R., Corbeil, J., van der Goot, F.G., and Aroian, R.V. (2004). Mitogen-activated protein kinase pathways defend against bacterial pore-forming toxins. Proc. Natl. Acad. Sci. U.S.A. 101, 10995–11000. Abstract Article
Hunter, C.P., and Kenyon, C. (1996). Spatial and temporal controls target pal-1 blastomere-specification activity to a single blastomere lineage in C. elegans embryos. Cell 87, 217–226. Abstract Article
Inoue, H., Hisamoto, N., An, J.H., Oliveira, R.P., Nishida, E., Blackwell, T.K., and Matsumoto, K. (2005). The C. elegans p38 MAPK pathway regulates nuclear localization of the transcription factor SKN-1 in oxidative stress response. Genes Dev. 19, 2278–2283. Abstract Article
Jee, C., Vanoaica, L., Lee, J., Park, B.J., and Ahnn, J. (2005). Thioredoxin is related to life span regulation and oxidative stress response in Caenorhabditis elegans. Genes Cells 10, 1203–1210. Abstract Article
Joshua, G.W. (2001). Functional analysis of leucine aminopeptidase in Caenorhabditis elegans. Mol. Biochem. Parasitol. 113, 223–232. Abstract Article
Kalb, J.M., Lau, K.K., Goszczynski, B., Fukushige, T., Moons, D., Okkema, P.G., and McGhee, J.D. (1998). pha-4 is Ce-fkh-1, a fork head/HNF-3 homolog that functions in organogenesis of the C. elegans pharynx. Development 125, 2171–2180. Abstract
Kang, S.H., and Kramer, J.M. (2000). Nidogen is nonessential and not required for normal type IV collagen localization in Caenorhabditis elegans. Mol. Biol. Cell 11, 3911–3923. Abstract Article
Kao, G., Huang, C.C., Hedgecock, E.M., Hall, D.H., and Wadsworth, W.G. (2006). The role of the laminin beta subunit in laminin heterotrimer assembly and basement membrane function and development in C. elegans. Dev. Biol. 290, 211–219. Abstract Article
Karabinos, A., Schmidt, H., Harborth, J., Schnabel, R., and Weber, K. (2001). Essential roles for four cytoplasmic intermediate filament proteins in Caenorhabditis elegans development. Proc. Natl. Acad. Sci. U.S.A. 98, 7863–7868. Abstract Article
Karabinos, A., Schulze, E., Klisch, T., Wang, J., and Weber, K. (2002). Expression profiles of the essential intermediate filament (IF) protein A2 and the IF protein C2 in the nematode Caenorhabditis elegans. Mech. Dev. 117, 311–314. Abstract Article
Karabinos, A., Bussing, I., Schulze, E., Wang, J., Weber, K., and Schnabel, R. (2003a). Functional analysis of the single calmodulin gene in the nematode Caenorhabditis elegans by RNA interference and 4-D microscopy. Eur. J. Cell. Biol. 82, 557–563. Abstract Article
Karabinos, A., Schulze, E., Schunemann, J., Parry, D.A., and Weber, K. (2003b). In vivo and in vitro evidence that the four essential intermediate filament (IF) proteins A1, A2, A3 and B1 of the nematode Caenorhabditis elegans form an obligate heteropolymeric IF system. J. Mol. Biol. 333, 307–319. Abstract Article
Karabinos, A., Schunemann, J., and Weber, K. (2004). Most genes encoding cytoplasmic intermediate filament (IF) proteins of the nematode Caenorhabditis elegans are required in late embryogenesis. Eur. J. Cell. Biol. 83, 457–468. Abstract Article
Katsura, I., Kondo, K., Amano, T., Ishihara, T., and Kawakami, M. (1994). Isolation, characterization and epistasis of fluoride-resistant mutants of Caenorhabditis elegans. Genetics 136, 145–154. Abstract
Kennedy, B.P., Aamodt, E.J., Allen, F.L., Chung, M.A., Heschl, M.F., and McGhee, J.D. (1993). The gut esterase gene (ges-1) from the nematodes Caenorhabditis elegans and Caenorhabditis briggsae. J. Mol. Biol. 229, 890–908. Abstract Article
Kim, Y.I., Cho, J.H., Yoo, O.J., and Ahnn, J. (2004). Transcriptional regulation and life-span modulation of cytosolic aconitase and ferritin genes in C. elegans. J. Mol. Biol. 342, 421–433. Abstract Article
Kimble, J., and Sharrock, W.J. (1983). Tissue-specific synthesis of yolk proteins in Caenorhabditis elegans. Dev. Biol. 96, 189–196. Abstract Article
Kipreos, E.T. (2005). C. elegans cell cycles: invariance and stem cell divisions. Nat. Rev. Mol. Cell Biol. 6, 766–776. Abstract Article
Kniazeva, M., Crawford, Q.T., Seiber, M., Wang, C.Y., and Han, M. (2004). Monomethyl branched-chain fatty acids play an essential role in Caenorhabditis elegans development. PLoS Biol. 2, E257. Abstract Article
Kniazeva, M., Sieber, M., McCauley, S., Zhang, K., Watts, J.L., and Han, M. (2003). Suppression of the ELO-2 FA elongation activity results in alterations of the fatty acid composition and multiple physiological defects, including abnormal ultradian rhythms, in Caenorhabditis elegans. Genetics 163, 159–169. Abstract
Koreth, J., and van den Heuvel, S. (2005). Cell-cycle control in Caenorhabditis elegans: how the worm moves from G1 to S. Oncogene 24, 2756–2764. Abstract Article
Kormish, J.D., and McGhee, J.D. (2005). The C. elegans lethal gut-obstructed gob-1 gene is trehalose-6-phosphate phosphatase. Dev. Biol. 26, 26. Abstract Article
Kostic, I., and Roy, R. (2002). Organ-specific cell division abnormalities caused by mutation in a general cell cycle regulator in C. elegans. Development 129, 2155–2165. Abstract
Kostich, M., Fire, A., and Fambrough, D.M. (2000). Identification and molecular-genetic characterization of a LAMP/CD68-like protein from Caenorhabditis elegans. J. Cell Sci. 113, 2595–2606. Abstract
Labrousse, A.M., Shurland, D.L., and van der Bliek, A.M. (1998). Contribution of the GTPase domain to the subcellular localization of dynamin in the nematode Caenorhabditis elegans. Mol. Biol. Cell 9, 3227–3239. Abstract
Lamitina, S.T., and Strange, K. (2005). Transcriptional targets of DAF-16 insulin signaling pathway protect C. elegans from extreme hypertonic stress. Am. J. Physiol. Cell Physiol. 288, C467–474. Abstract Article
Laurent, V., Brooks, D.R., Coates, D., and Isaac, R.E. (2001). Functional expression and characterization of the cytoplasmic aminopeptidase P of Caenorhabditis elegans. Eur. J. Biochem. 268, 5430–5438. Abstract Article
Legouis, R., Gansmuller, A., Sookhareea, S., Bosher, J.M., Baillie, D.L., and Labouesse, M. (2000). LET-413 is a basolateral protein required for the assembly of adherens junctions in Caenorhabditis elegans. Nat. Cell Biol. 2, 415–422. Abstract Article
Legouis, R., Jaulin-Bastard, F., Schott, S., Navarro, C., Borg, J.P., and Labouesse, M. (2003). Basolateral targeting by leucine-rich repeat domains in epithelial cells. EMBO Rep. 4, 1096–1102. Abstract Article
Lehane, M.J. (1997). Peritrophic matrix structure and function. Annu. Rev. Entomol. 42, 525–550. Abstract Article
Leung, B., Hermann, G.J., and Priess, J.R. (1999). Organogenesis of the Caenorhabditis elegans intestine. Dev. Biol. 216, 114–134. Abstract Article
Liao, V.H., Dong, J., and Freedman, J.H. (2002). Molecular characterization of a novel, cadmium-inducible gene from the nematode Caenorhabditis elegans. A new gene that contributes to the resistance to cadmium toxicity. J. Biol. Chem. 277, 42049–42059. Abstract Article
Liao, V.H., and Freedman, J.H. (1998). Cadmium-regulated genes from the nematode Caenorhabditis elegans. Identification and cloning of new cadmium-responsive genes by differential display. J. Biol. Chem. 273, 31962–31970. Abstract Article
Libina, N., Berman, J.R., and Kenyon, C. (2003). Tissue-specific activities of C. elegans DAF-16 in the regulation of lifespan. Cell 115, 489–502. Abstract Article
Lin, K., Hsin, H., Libina, N., and Kenyon, C. (2001). Regulation of the Caenorhabditis elegans longevity protein DAF-16 by insulin/IGF-1 and germline signaling. Nat. Genet. 28, 139–145. Abstract Article
Lin, R., Hill, R.J., and Priess, J.R. (1998). POP-1 and anterior-posterior fate decisions in C. elegans embryos. Cell 92, 229–239. Abstract Article
Lin, R., Thompson, S., and Priess, J.R. (1995). pop-1 encodes an Hmg box protein required for the specification of a mesoderm precursor in early C. elegans embryos. Cell 83, 599–609. Abstract Article
Lincke, C.R., Broeks, A., The, I., Plasterk, R.H., and Borst, P. (1993). The expression of two P-glycoprotein (pgp) genes in transgenic Caenorhabditis elegans is confined to intestinal cells. EMBO J. 12, 1615–1620. Abstract
Lindblom, T.H., Pierce, G.J., and Sluder, A.E. (2001). A C. elegans orphan nuclear receptor contributes to xenobiotic resistance. Curr. Biol. 11, 864–868. Abstract Article
Lo, M.C., Gay, F., Odom, R., Shi, Y., and Lin, R. (2004). Phosphorylation by the beta-catenin/MAPK complex promotes 14-3-3-mediated nuclear export of TCF/POP-1 in signal-responsive cells in C. elegans. Cell 117, 95–106. Abstract Article
Luersen, K., Eschbach, M.L., Liebau, E., and Walter, R.D. (2004). Functional GATA- and initiator-like-elements exhibit a similar arrangement in the promoters of Caenorhabditis elegans polyamine synthesis enzymes. Biol. Chem. 385, 711–721. Abstract Article
MacMorris, M., and Blumenthal, T. (1993). In situ analysis of C. elegans vitellogenin fusion gene expression in integrated transgenic strains: effect of promoter mutations on RNA localization. Gene Expr. 3, 27–36. Abstract
MacMorris, M., Broverman, S., Greenspoon, S., Lea, K., Madej, C., Blumenthal, T., and Spieth, J. (1992). Regulation of vitellogenin gene expression in transgenic Caenorhabditis elegans: short sequences required for activation of the vit-2 promoter. Mol. Cell. Biol. 12, 1652–1662. Abstract
MacMorris, M., Spieth, J., Madej, C., Lea, K., and Blumenthal, T. (1994). Analysis of the VPE sequences in the Caenorhabditis elegans vit-2 promoter with extrachromosomal tandem array-containing transgenic strains. Mol. Cell. Biol. 14, 484–491. Abstract
MacQueen, A.J., Baggett, J.J., Perumov, N., Bauer, R.A., Januszewski, T., Schriefer, L., and Waddle, J.A. (2005). ACT-5 is an essential Caenorhabditis elegans actin required for intestinal microvilli formation. Mol. Biol. Cell 16, 3247–3259. Abstract Article
Maduro, M.F., Broitman-Maduro, G., Mengarelli, I., and Rothman, J.H. (2006). Maternal deployment of the embryonic SKN-1 MED-1,2 cell specification pathway in C. elegans. Dev Biol. Abstract Article
Maduro, M.F., Hill, R.J., Heid, P.J., Newman-Smith, E.D., Zhu, J., Priess, J.R., and Rothman, J.H. (2005). Genetic redundancy in endoderm specification within the genus Caenorhabditis. Dev. Biol. 284, 509–522. Abstract Article
Maduro, M.F., Kasmir, J.J., Zhu, J., and Rothman, J.H. (2005). The Wnt effector POP-1 and the PAL-1/Caudal homeoprotein collaborate with SKN-1 to activate C. elegans endoderm development. Dev. Biol. 285, 510–523. Abstract Article
Maduro, M.F., Meneghini, M.D., Bowerman, B., Broitman-Maduro, G., and Rothman, J.H. (2001). Restriction of mesendoderm to a single blastomere by the combined action of SKN-1 and a GSK-3beta homolog is mediated by MED-1 and -2 in C. elegans. Mol. Cell 7, 475–485. Abstract Article
Maduro, M.F., and Rothman, J.H. (2002). Making worm guts: the gene regulatory network of the Caenorhabditis elegans endoderm. Dev. Biol. 246, 68–85. Abstract Article
Maebuchi, M., Togo, S.H., Yokota, S., Ghenea, S., Bun-Ya, M., Kamiryo, T., and Kawahara, A. (1999). Type-II 3-oxoacyl-CoA thiolase of the nematode Caenorhabditis elegans is located in peroxisomes, highly expressed during larval stages and induced by clofibrate. Eur. J. Biochem. 264, 509–515. Abstract Article
Mak, H.Y., Nelson, L.S., Basson, M., Johnson, C.D., and Ruvkun, G. (2006). Polygenic control of Caenorhabditis elegans fat storage. Nat. Genet. 38, 363–368. Abstract Article
Mallo, G.V., Kurz, C.L., Couillault, C., Pujol, N., Granjeaud, S., Kohara, Y., and Ewbank, J.J. (2002). Inducible antibacterial defense system in C. elegans. Curr. Biol. 12, 1209–1214. Abstract Article
Mango, S.E., Lambie, E.J., and Kimble, J. (1994). The pha-4 gene is required to generate the pharyngeal primordium of Caenorhabditis elegans. Development 120, 3019–3031. Abstract
Marroquin, L.D., Elyassnia, D., Griffitts, J.S., Feitelson, J.S., and Aroian, R.V. (2000). Bacillus thuringiensis (Bt) toxin susceptibility and isolation of resistance mutants in the nematode Caenorhabditis elegans. Genetics 155, 1693–1699. Abstract
Marshall, S.D., and McGhee, J.D. (2001). Coordination of ges-1 expression between the Caenorhabditis pharynx and intestine. Dev. Biol. 239, 350–363. Abstract Article
McElwee, J., Bubb, K., and Thomas, J.H. (2003). Transcriptional outputs of the Caenorhabditis elegans forkhead protein DAF-16. Aging Cell 2, 111–121. Abstract Article
McGhee, J.D. (1987). Purification and characterization of a carboxylesterase from the intestine of the nematode Caenorhabditis elegans. Biochemistry 26, 4101–4107. Abstract Article
McGhee, J.D., Sleumer, M.C., Bilenky, M., Wong, K., McKay, S.J., Goszczynski, B., Tian, H., Krich, N., Khattra, J., Holt, R.A., et al. (2007). The ELT-2 GATA-Factor and the Global Regulation of Transcription in the C. elegans Intestine. Dev. Biol. 302, 627–645. Abstract Article
McKay, R.M., McKay, J.P., Avery, L., and Graff, J.M. (2003). C. elegans: a model for exploring the genetics of fat storage. Dev. Cell 4, 131–142. Abstract Article
McMahon, L., Legouis, R., Vonesch, J.L., and Labouesse, M. (2001). Assembly of C. elegans apical junctions involves positioning and compaction by LET-413 and protein aggregation by the MAGUK protein DLG-1. J. Cell Sci. 114, 2265–2277. Abstract Article
Meissner, B., Boll, M., Daniel, H., and Baumeister, R. (2004). Deletion of the intestinal peptide transporter affects insulin and TOR signaling in Caenorhabditis elegans. J. Biol. Chem. 279, 36739–36745. Abstract Article
Melendez, A., Talloczy, Z., Seaman, M., Eskelinen, E.L., Hall, D.H., and Levine, B. (2003). Autophagy genes are essential for dauer development and life-span extension in C. elegans. Science 301, 1387–1391. Abstract Article
Mendel, J., Heinecke, K., Fyrst, H., and Saba, J.D. (2003). Sphingosine phosphate lyase expression is essential for normal development in Caenorhabditis elegans. J. Biol. Chem. 278, 22341–22349. Abstract Article
Menzel, R., Bogaert, T., and Achazi, R. (2001). A systematic gene expression screen of Caenorhabditis elegans cytochrome P450 genes reveals CYP35 as strongly xenobiotic inducible. Arch. Biochem. Biophys. 395, 158–168. Abstract Article
Miller, K.G., Emerson, M.D., and Rand, J.B. (1999). Goalpha and diacylglycerol kinase negatively regulate the Gqalpha pathway in C. elegans. Neuron 24, 323–333. Abstract Article
Moerman, D.G., Hutter, H., Mullen, G.P., and Schnabel, R. (1996). Cell autonomous expression of perlecan and plasticity of cell shape in embryonic muscle of Caenorhabditis elegans. Dev. Biol. 173, 228–242. Abstract Article
Moilanen, L.H., Fukushige, T., and Freedman, J.H. (1999). Regulation of metallothionein gene transcription. Identification of upstream regulatory elements and transcription factors responsible for cell-specific expression of the metallothionein genes from Caenorhabditis elegans. J. Biol. Chem. 274, 29655–29665. Abstract Article
Munn, E., and Greenwood, C. (1984). The occurrence of submicrovillar endotube (Modified Terminal Web) and associated cytoskeletal structures in the intestinal epithelia of nematodes. Philos. Trans. R. Soc. Lond., B, Biol. Sci. 306, 1–18. Article
Munn, E.A., and Munn, P.D. (2002). Feeding and Digestion, In Biology of Nematodes, D.L. Lee, ed. (London: Taylor&Francis).
Murphy, C.T., McCarroll, S.A., Bargmann, C.I., Fraser, A., Kamath, R.S., Ahringer, J., Li, H., and Kenyon, C. (2003). Genes that act downstream of DAF-16 to influence the lifespan of Caenorhabditis elegans. Nature 424, 277–283. Abstract Article
Nash, B., Colavita, A., Zheng, H., Roy, P.J., and Culotti, J.G. (2000). The forkhead transcription factor UNC-130 is required for the graded spatial expression of the UNC-129 TGF-beta guidance factor in C. elegans. Genes Dev. 14, 2486–2500. Abstract Article
Nehrke, K. (2003). A reduction in intestinal cell pHi due to loss of the Caenorhabditis elegans Na+/H+ exchanger NHX-2 increases life span. J. Biol. Chem. 278, 44657–44666. Abstract Article
Nehrke, K., and Melvin, J.E. (2002). The NHX family of Na+-H+ exchangers in Caenorhabditis elegans. J. Biol. Chem. 277, 29036–29044. Abstract Article
Neves, A., and Priess, J.R. (2005). The REF-1 family of bHLH transcription factors pattern C. elegans embryos through Notch-dependent and Notch-independent pathways. Dev. Cell 8, 867–879. Abstract Article
Norman, K.R., and Moerman, D.G. (2000). The let-268 locus of Caenorhabditis elegans encodes a procollagen lysyl hydroxylase that is essential for type IV collagen secretion. Dev. Biol. 227, 690–705. Abstract Article
Oka, T., and Futai, M. (2000). Requirement of V-ATPase for ovulation and embryogenesis in Caenorhabditis elegans. J. Biol. Chem. 275, 29556–29561. Abstract Article
Oka, T., Toyomura, T., Honjo, K., Wada, Y., and Futai, M. (2001). Four subunit a isoforms of Caenorhabditis elegans vacuolar H+-ATPase. Cell-specific expression during development. J. Biol. Chem. 276, 33079–33085. Abstract Article
Oskouian, B., Mendel, J., Shocron, E., Lee, M.A., Jr., Fyrst, H., and Saba, J.D. (2005). Regulation of sphingosine-1-phosphate lyase gene expression by members of the GATA family of transcription factors. J. Biol. Chem. 280, 18403–18410. Abstract Article
Park, B.J., Lee, D.G., Yu, J.R., Jung, S.K., Choi, K., Lee, J., Kim, Y.S., Lee, J.I., Kwon, J.Y., Singson, A., et al. (2001). Calreticulin, a calcium-binding molecular chaperone, is required for stress response and fertility in Caenorhabditis elegans. Mol. Biol. Cell 12, 2835–2845. Abstract
Pauli, F., Liu, Y., Kim, Y.A., Chen, P.J., and Kim, S.K. (2006). Chromosomal clustering and GATA transcriptional regulation of intestine-expressed genes in C. elegans. Development 133, 287–295. Abstract Article
Pocock, R., Ahringer, J., Mitsch, M., Maxwell, S., and Woollard, A. (2004). A regulatory network of T-box genes and the even-skipped homologue vab-7 controls patterning and morphogenesis in C. elegans. Development 131, 2373–2385. Abstract Article
Popham, J., and Webster, J. (1979). Aspects of the fine structure of the dauer larva of the nematode Caenorhabditis elegans. Can. J. Zool. 57, 794–800.
Poulin, G., Dong, Y., Fraser, A.G., Hopper, N.A., and Ahringer, J. (2005). Chromatin regulation and sumoylation in the inhibition of Ras-induced vulval development in Caenorhabditis elegans. EMBO J. 24, 2613–2623. Abstract Article
Raymond, C.S., Shamu, C.E., Shen, M.M., Seifert, K.J., Hirsch, B., Hodgkin, J., and Zarkower, D. (1998). Evidence for evolutionary conservation of sex-determining genes. Nature 391, 691–695. Abstract Article
Robinson-Rechavi, M., Maina, C.V., Gissendanner, C.R., Laudet, V., and Sluder, A. (2005). Explosive lineage-specific expansion of the orphan nuclear receptor HNF4 in nematodes. J. Mol. Evol. 60, 577–586. Abstract Article
Rocheleau, C.E., Downs, W.D., Lin, R., Wittmann, C., Bei, Y., Cha, Y.H., Ali, M., Priess, J.R., and Mello, C.C. (1997). Wnt signaling and an APC-related gene specify endoderm in early C. elegans embryos. Cell 90, 707–716. Abstract Article
Rolls, M.M., Hall, D.H., Victor, M., Stelzer, E.H., and Rapoport, T.A. (2002). Targeting of rough endoplasmic reticulum membrane proteins and ribosomes in invertebrate neurons. Mol. Biol. Cell 13, 1778–1791. Abstract Article
Ross, J.M., and Zarkower, D. (2003). Polycomb group regulation of Hox gene expression in C. elegans. Dev. Cell 4, 891–901. Abstract Article
Rottiers, V., Motola, D.L., Gerisch, B., Cummins, C.L., Nishiwaki, K., Mangelsdorf, D.J., and Antebi, A. (2006). Hormonal control of C. elegans dauer formation and life span by a Rieske-like oxygenase. Dev. Cell 10, 473–482. Abstract Article
Roudier, N., Lefebvre, C., and Legouis, R. (2005). CeVPS-27 is an endosomal protein required for the molting and the endocytic trafficking of the low-density lipoprotein receptor-related protein 1 in Caenorhabditis elegans. Traffic 6, 695–705. Abstract Article
Ruvinsky, I., and Ruvkun, G. (2003). Functional tests of enhancer conservation between distantly related species. Development 130, 5133–5142. Abstract Article
Saifee, O., Wei, L., and Nonet, M.L. (1998). The Caenorhabditis elegans unc-64 locus encodes a syntaxin that interacts genetically with synaptobrevin. Mol. Biol. Cell 9, 1235–1252. Abstract
Sarkis, G.J., Kurpiewski, M.R., Ashcom, J.D., Jen-Jacobson, L., and Jacobson, L.A. (1988). Proteases of the nematode Caenorhabditis elegans. Arch. Biochem. Biophys. 261, 80–90. Abstract Article
Sato, M., Sato, K., Fonarev, P., Huang, C.J., Liou, W., and Grant, B.D. (2005). Caenorhabditis elegans RME-6 is a novel regulator of RAB-5 at the clathrin-coated pit. Nat. Cell Biol. 7, 559–569. Abstract Article
Schimpf, J., Sames, K., and Zwilling, R. (1999). Proteoglycan distribution pattern during aging in the nematode Caenorhabditis elegans: an ultrastructural histochemical study. Histochem. J. 31, 285–292. Abstract Article
Schroeder, D.F., and McGhee, J.D. (1998). Anterior-posterior patterning within the Caenorhabditis elegans endoderm. Development 125, 4877–4887. Abstract
Schulenburg, H., Kurz, C.L., and Ewbank, J.J. (2004). Evolution of the innate immune system: the worm perspective. Immunol. Rev. 198, 36–58. Abstract Article
Segbert, C., Johnson, K., Theres, C., van Furden, D., and Bossinger, O. (2004). Molecular and functional analysis of apical junction formation in the gut epithelium of Caenorhabditis elegans. Dev. Biol. 266, 17–26. Abstract Article
Shen, M.M., and Hodgkin, J. (1988). mab-3, a gene required for sex-specific yolk protein expression and a male-specific lineage in C. elegans. Cell 54, 1019–1031. Abstract Article
Sherman, T., Chernova, M.N., Clark, J.S., Jiang, L., Alper, S.L., and Nehrke, K. (2005). The abts and sulp families of anion transporters from Caenorhabditis elegans. Am. J. Physiol. Cell Physiol. 289, C341–351. Abstract Article
Shetty, P., Lo, M.C., Robertson, S.M., and Lin, R. (2005). C. elegans TCF protein, POP-1, converts from repressor to activator as a result of Wnt-induced lowering of nuclear levels. Dev. Biol. 285, 584–592. Abstract Article
Simske, J.S., Kaech, S.M., Harp, S.A., and Kim, S.K. (1996). LET-23 receptor localization by the cell junction protein LIN-7 during C. elegans vulval induction. Cell 85, 195–204. Abstract Article
Slice, L.W., Freedman, J.H., and Rubin, C.S. (1990). Purification, characterization, and cDNA cloning of a novel metallothionein-like, cadmium-binding protein from Caenorhabditis elegans. J. Biol. Chem. 265, 256–263. Abstract
Spieth, J., Shim, Y.H., Lea, K., Conrad, R., and Blumenthal, T. (1991). elt-1, an embryonically expressed Caenorhabditis elegans gene homologous to the Gata transcription factor family. Mol. Cell. Biol. 11, 4651–4659. Abstract
Starich, T.A., Miller, A., Nguyen, R.L., Hall, D.H., and Shaw, J.E. (2003). The Caenorhabditis elegans innexin INX-3 is localized to gap junctions and is essential for embryonic development. Dev. Biol. 256, 403–417. Abstract Article
Starr, D.A., Hermann, G.J., Malone, C.J., Fixsen, W., Priess, J.R., Horvitz, H.R., and Han, M. (2001). unc-83 encodes a novel component of the nuclear envelope and is essential for proper nuclear migration. Development 128, 5039–5050. Abstract
Sulston, J.E. (1976). Post-embryonic development in the ventral cord of Caenorhabditis elegans. Philos. Trans. R. Soc. Lond., B, Biol. Sci. 275, 287–297. Abstract
Sulston, J.E., and Horvitz, H.R. (1977). Post-embryonic cell lineages of the nematode, Caenorhabditis elegans. Dev. Biol. 56, 110–156. Abstract Article
Sulston, J.E., Schierenberg, E., White, J.G., and Thomson, J.N. (1983). The embryonic cell lineage of the nematode Caenorhabditis elegans. Dev. Biol. 100, 64–119. Abstract Article
Sundaram, P., Echalier, B., Han, W., Hull, D., and Timmons, L. (2006). ATP-binding cassette transporters are required for efficient RNA interference in Caenorhabditis elegans. Mol. Biol. Cell 17, 3678–3688. Abstract Article
Swain, S.C., Keusekotten, K., Baumeister, R., and Sturzenbaum, S.R. (2004). C. elegans metallothioneins: new insights into the phenotypic effects of cadmium toxicosis. J. Mol. Biol. 341, 951–959. Abstract Article
Take-Uchi, M., Kawakami, M., Ishihara, T., Amano, T., Kondo, K., and Katsura, I. (1998). An ion channel of the degenerin/epithelial sodium channel superfamily controls the defecation rhythm in Caenorhabditis elegans. Proc. Natl. Acad. Sci. U.S.A. 95, 11775–11780. Abstract Article
Take-Uchi, M., Kobayashi, Y., Kimura, K.D., Ishihara, T., and Katsura, I. (2005). FLR-4, a Novel Serine/Threonine Protein Kinase, Regulates Defecation Rhythm in Caenorhabditis elegans. Mol. Biol. Cell 16, 1355–1365. Abstract Article
Taubert, S., Van Gilst, M.R., Hansen, M., and Yamamoto, K.R. (2006). A Mediator subunit, MDT-15, integrates regulation of fatty acid metabolism by NHR-49-dependent and -independent pathways in C. elegans. Genes Dev. 20, 1137–1149. Abstract Article
Tcherepanova, I., Bhattacharyya, L., Rubin, C.S., and Freedman, J.H. (2000). Aspartic proteases from the nematode Caenorhabditis elegans. Structural organization and developmental and cell-specific expression of asp-1. J. Biol. Chem. 275, 26359–26369. Abstract Article
Tenenhaus, C., Subramaniam, K., Dunn, M.A., and Seydoux, G. (2001). PIE-1 is a bifunctional protein that regulates maternal and zygotic gene expression in the embryonic germ line of Caenorhabditis elegans. Genes Dev. 15, 1031–1040. Abstract Article
Teramoto, T., Lambie, E.J., and Iwasaki, K. (2005). Differential regulation of TRPM channels governs electrolyte homeostasis in the C. elegans intestine. Cell Metab 1, 343–354. Abstract Article
Teramoto, T., and Iwasaki, K. (2006). Intestinal calcium waves coordinate a behavioral motor program in C. elegans. Cell Calcium 40, 319–327. Abstract Article
Thieringer, H., Moellers, B., Dodt, G., Kunau, W.H., and Driscoll, M. (2003). Modeling human peroxisome biogenesis disorders in the nematode Caenorhabditis elegans. J. Cell Sci. 116, 1797–1804. Abstract Article
Thompson, F.J., Britton, C., Wheatley, I., Maitland, K., Walker, G., Anant, S., Davidson, N.O., and Devaney, E. (2002). Biochemical and molecular characterization of two cytidine deaminases in the nematode Caenorhabditis elegans. Biochem. J. 365, 99–107. Abstract Article
Thorpe, C.J., Schlesinger, A., Carter, J.C., and Bowerman, B. (1997). Wnt signaling polarizes an early C. elegans blastomere to distinguish endoderm from mesoderm. Cell 90, 695–705. Abstract Article
Togo, S.H., Maebuchi, M., Yokota, S., Bun-Ya, M., Kawahara, A., and Kamiryo, T. (2000). Immunological detection of alkaline-diaminobenzidine-negative peroxisomes of the nematode Caenorhabditis elegans. Eur. J. Biochem. 267, 1307–1312. Abstract Article
Van Auken, K., Weaver, D., Robertson, B., Sundaram, M., Saldi, T., Edgar, L., Elling, U., Lee, M., Boese, Q., and Wood, W.B. (2002). Roles of the Homothorax/Meis/Prep homolog UNC-62 and the Exd/Pbx homologs CEH-20 and CEH-40 in C. elegans embryogenesis. Development 129, 5255–5268. Abstract
Van Furden, D., Johnson, K., Segbert, C., and Bossinger, O. (2004). The C. elegans ezrin-radixin-moesin protein ERM-1 is necessary for apical junction remodelling and tubulogenesis in the intestine. Dev. Biol. 272, 262–276. Abstract Article
Vogel, B.E., and Hedgecock, E.M. (2001). Hemicentin, a conserved extracellular member of the immunoglobulin superfamily, organizes epithelial and other cell attachments into oriented line-shaped junctions. Development 128, 883–894. Abstract
Walker, D.S., Ly, S., Gower, N.J., and Baylis, H.A. (2004). IRI-1, a LIN-15B homologue, interacts with inositol-1,4,5-triphosphate receptors and regulates gonadogenesis, defecation, and pharyngeal pumping in Caenorhabditis elegans. Mol. Biol. Cell 15, 3073–3082. Abstract Article
Walker, G., Houthoofd, K., Vanfleteren, J.R., and Gems, D. (2005). Dietary restriction in C. elegans: from rate-of-living effects to nutrient sensing pathways. Mech. Ageing Dev. 126, 929–937. Abstract Article
Warren, C.E., Krizus, A., and Dennis, J.W. (2001). Complementary expression patterns of six nonessential Caenorhabditis elegans core 2/I N-acetylglucosaminyltransferase homologues. Glycobiology 11, 979–988. Abstract Article
Watts, J.L., and Browse, J. (2006). Dietary manipulation implicates lipid signaling in the regulation of germ cell maintenance in C. elegans. Dev. Biol. 292, 381–392. Abstract Article
White, J. (1988). The Anatomy, In The Nematode Caenorhabditis elegans, W.B. Wood, ed. (Plainview, New York: Cold Spring Harbor Laboratory Press), pp. 81–122.
Winston, W.M., Molodowitch, C., and Hunter, C.P. (2002a). Systemic RNAi in C. elegans requires the putative transmembrane protein SID-1. Science 295, 2456–2459. Abstract Article
Winston, W.M., Molodowitch, C., and Hunter, C.P. (2002b). Systemic RNAi in C. elegans requires the putative transmembrane protein SID-1. Science 295, 2456–2459. Abstract Article
Wittmann, C., Bossinger, O., Goldstein, B., Fleischmann, M., Kohler, R., Brunschwig, K., Tobler, H., and Muller, F. (1997). The expression of the C. elegans labial-like Hox gene ceh-13 during early embryogenesis relies on cell fate and on anteroposterior cell polarity. Development 124, 4193–4200. Abstract Article
Wood, W.B. (1991). Evidence from reversal of handedness in C. elegans embryos for early cell interactions determining cell fates. Nature 349, 536–538. Abstract Article
Wood, W.B., Bergmann, D., and Florance, A. (1996). Maternal effect of low temperature on handedness determination in C. elegans embryos. Dev. Genet. 19, 222–230. Abstract
Wood, W.B., Hecht, R., Carr, S., Vanderslice, R., Wolf, N., and Hirsh, D. (1980). Parental effects and phenotypic characterization of mutations that affect early development in Caenorhabditis elegans. Dev. Biol. 74, 446–469. Abstract Article
Wu, S.L., Staudinger, J., Olson, E.N., and Rubin, C.S. (1998). Structure, expression, and properties of an atypical protein kinase C (PKC3) from Caenorhabditis elegans. PKC3 is required for the normal progression of embryogenesis and viability of the organism. J. Biol. Chem. 273, 1130–1143. Abstract Article
Wu, Y.C., Stanfield, G.M., and Horvitz, H.R. (2000). NUC-1, a Caenorhabditis elegans DNase II homolog, functions in an intermediate step of DNA degradation during apoptosis. Genes Dev. 14, 536–548. Abstract
Xue, Y., Fares, H., Grant, B., Li, Z., Rose, A.M., Clark, S.G., and Skolnik, E.Y. (2003). Genetic analysis of the myotubularin family of phosphatases in Caenorhabditis elegans. J. Biol. Chem. 278, 34380–34386. Abstract Article
Yamanaka, A., Yada, M., Imaki, H., Koga, M., Ohshima, Y., and Nakayama, K. (2002). Multiple Skp1-related proteins in Caenorhabditis elegans: diverse patterns of interaction with Cullins and F-box proteins. Curr. Biol. 12, 267–275. Abstract Article
Yang, F., Vought, B.W., Satterlee, J.S., Walker, A.K., Jim Sun, Z.Y., Watts, J.L., DeBeaumont, R., Saito, R.M., Hyberts, S.G., Yang, S., et al. (2006). An ARC/Mediator subunit required for SREBP control of cholesterol and lipid homeostasis. Nature 442, 700–704. Abstract Article
Yi, W., and Zarkower, D. (1999). Similarity of DNA binding and transcriptional regulation by Caenorhabditis elegans MAB-3 and Drosophila melanogaster DSX suggests conservation of sex determining mechanisms. Development 126, 873–881. Abstract
Yokota, S., Togo, S.H., Maebuchi, M., Bun-Ya, M., Haraguchi, C.M., and Kamiryo, T. (2002). Peroxisomes of the nematode Caenorhabditis elegans: distribution and morphological characteristics. Histochem. Cell Biol. 118, 329–336. Abstract Article
Yu, X., Odera, S., Chuang, C.H., Lu, N., and Zhou, Z. (2006). C. elegans Dynamin mediates the signaling of phagocytic receptor CED-1 for the engulfment and degradation of apoptotic cells. Dev. Cell 10, 743–757. Abstract Article
Zhang, L., Wu, S.L., and Rubin, C.S. (2001). A novel adapter protein employs a phosphotyrosine binding domain and exceptionally basic N-terminal domains to capture and localize an atypical protein kinase C: characterization of Caenorhabditis elegans C kinase adapter 1, a protein that avidly binds protein kinase C3. J. Biol. Chem. 276, 10463–10475. Abstract Article
Zheng, Q., Van Die, I., and Cummings, R.D. (2002). Molecular cloning and characterization of a novel alpha 1,2-fucosyltransferase (CE2FT-1) from Caenorhabditis elegans. J. Biol. Chem. 277, 39823–39832. Abstract Article
Zhu, J., Fukushige, T., McGhee, J.D., and Rothman, J.H. (1998). Reprogramming of early embryonic blastomeres into endodermal progenitors by a Caenorhabditis elegans GATA factor. Genes Dev. 12, 3809–3814. Abstract
*Edited by Geraldine Seydoux and James R. Priess. Last revised December 13, 2006. Published March 27, 2007. This chapter should be cited as: McGhee, J.D. The C. elegans intestine (March 27, 2007), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.133.1, http://www.wormbook.org.
Copyright: © 2007 James D. McGhee. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
§To whom correspondence should be addressed. E-mail: jmcghee@ucalgary.ca
 All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.
All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.