Embryo series courtesy of Einhard Schierenberg
Embryo series courtesy of Einhard SchierenbergAbstract
MicroRNAs (miRNAs) are small, non-coding regulatory RNAs found in many phyla that control such diverse events as development, metabolism, cell fate and cell death. They have also been implicated in human cancers. The C. elegans genome encodes hundreds of miRNAs, including the founding members of the miRNA family lin-4 and let-7. Despite the abundance of C. elegans miRNAs, few miRNA targets are known and little is known about the mechanism by which they function. However, C. elegans research continues to push the boundaries of discovery in this area.
lin-4 and let-7 are the best understood miRNAs. They control the timing of adult cell fate determination in hypodermal cells by binding to partially complementary sites in the mRNA of key developmental regulators to repress protein expression. For example, lin-4 is predicted to bind to seven sites in the lin-14 3' untranslated region (UTR) to repress LIN-14, while let-7 is predicted to bind two let-7 complementary sites in the lin-41 3' UTR to down-regulate LIN-41. Two other miRNAs, lsy-6 and mir-273, control left-right asymmetry in neural development, and also target key developmental regulators for repression. Approximately one third of the C. elegans miRNAs are differentially expressed during development indicating a major role for miRNAs in C. elegans development. Given the remarkable conservation of developmental mechanism across phylogeny, many of the principles of miRNAs discovered in C. elegans are likely to be applicable to higher animals.
MicroRNAs (miRNAs) are genomically-encoded, untranslated RNA molecules of approximately 20-25 nucleotides (nt). MiRNAs regulate diverse processes such as cell fate specification, apoptosis, and metabolism (Bartel, 2004). In a few short years, these tiny RNAs have impacted virtually every field of biology and changed the way we view gene regulation. Victor Ambros and colleagues identified the first miRNA, lin-4, in C. elegans over a decade ago (Lee et al., 1993). The lin-4 gene was unusual in that it did not encode a protein but rather a small RNA that imperfectly base-paired to complementary sequences on target messenger RNAs (mRNAs) in order to block gene expression (Lee et al., 1993; Wightman et al., 1993). This prompted Wickens and Takayama to ask: "Is this regulatory RNA a deviant or an emissary?" (Wickens and Takayama, 1994). The answer is now clearly "emissary". In 2000, a second miRNA, let-7, was discovered by Gary Ruvkun's group to direct the later stages of C. elegans development in a similar manner to lin-4 (Reinhart et al., 2000). It was soon discovered that both lin-4 and let-7 were evolutionarily conserved from flies to humans, implicating a more universal role for these genes in animals (Lagos-Quintana et al., 2002; Pasquinelli et al., 2000). The miRNA field exploded after several laboratories revealed, using cloning and bioinformatics techniques, that there were hundreds of miRNAs present in both plant and animal genomes (Houbaviy et al., 2003; Lagos-Quintana et al., 2001; Lau et al., 2001; Lee and Ambros, 2001; Lim et al., 2003; Mourelatos et al., 2002; Reinhart et al., 2002). Like lin-4 and let-7, some of the recently discovered miRNAs are temporally regulated during development, while others are spatially expressed. Many are highly conserved in Drosophila and humans, suggesting ancient functions (Lau et al., 2001). Despite the large number of C. elegans miRNAs, the roles for most of them await discovery.
Generally, miRNAs are transcribed, some by RNA polyermase II (Bartel, 2004; Lee et al., 2004a), as longer poly-adenylylated primary miRNAs (pri-miRNAs) molecules of about 1 kilobase or greater (Bracht et al., 2004; Lee et al., 2002). The pri-miRNA is further processed by the RNase III endonuclease Drosha (drsh-1 in C. elegans; Bracht et al., 2004; Lee et al., 2003) to the precursor miRNA (pre-miRNA), a 60-70 nt molecule that can fold back on itself and form a hairpin loop structure. The pre-miRNA is then transported out of the nucleus into the cytoplasm by Exportin-5 (Yi et al., 2003). In the cytoplasm another RNase III enzyme, Dicer (dcr-1 in C. elegans), processes the pre-miRNA to the mature 20-25 nt miRNA (Bernstein et al., 2001; Grishok et al., 2001). Mature miRNAs bind to imperfectly complementary sequences in the 3' untranslated regions (UTRs) of target mRNAs to negatively regulate target gene expression. miRNAs are thus believed to function as guides to recruit a silencing complex to target mRNAs, but the exact mechanism used by miRNAs to down-regulate gene expression is unknown.
C. elegans has proven to be the pioneer organism for miRNA research. The C. elegans genome contains approximately 110 confirmed miRNA genes and estimates variously place the total as high as 300 (Grad et al., 2003; Lim et al., 2003). Approximately 34 C. elegans miRNAs are conserved in other phyla, (making up 22 families) and 30 are temporally regulated during development (Lim et al., 2003).
lin-4 and let-7, the first two known miRNAs, were discovered through conventional forward genetic screens in C. elegans for heterochronic (developmental timing) mutants (Chalfie et al., 1981; Lee et al., 1993; Reinhart et al., 2000). Briefly, lin-4 and let-7 are temporally regulated and govern temporal aspects of development in C. elegans (Feinbaum and Ambros, 1999; Johnson et al., 2003; Reinhart et al., 2000). These miRNAs down-regulate gene expression by interacting with partially complementary sequences in the 3' UTRs of their target genes, which are themselves important heterochronic genes. More recently, the role of two other C. elegans miRNAs have been discovered in spatial pattern formation in the nervous system (Chang et al., 2004; Johnston and Hobert, 2003), demonstrating the diversity of roles played by miRNAs. Much remains to be discovered about the biological roles of the other C. elegans miRNAs, but their sheer number suggests that their impact will be extensive.
lin-4 is required for a normal L1-to-L2 transition during C. elegans post-embryonic development (Ambros and Horvitz, 1984) and encodes at least two small RNA molecules, a rare 61 nt pre-lin-4 and a more abundant 22 nt mature lin-4 (Lee et al., 1993). lin-4 is related to another temporally-expressed C. elegans miRNA, mir-237 (Lim et al., 2003).
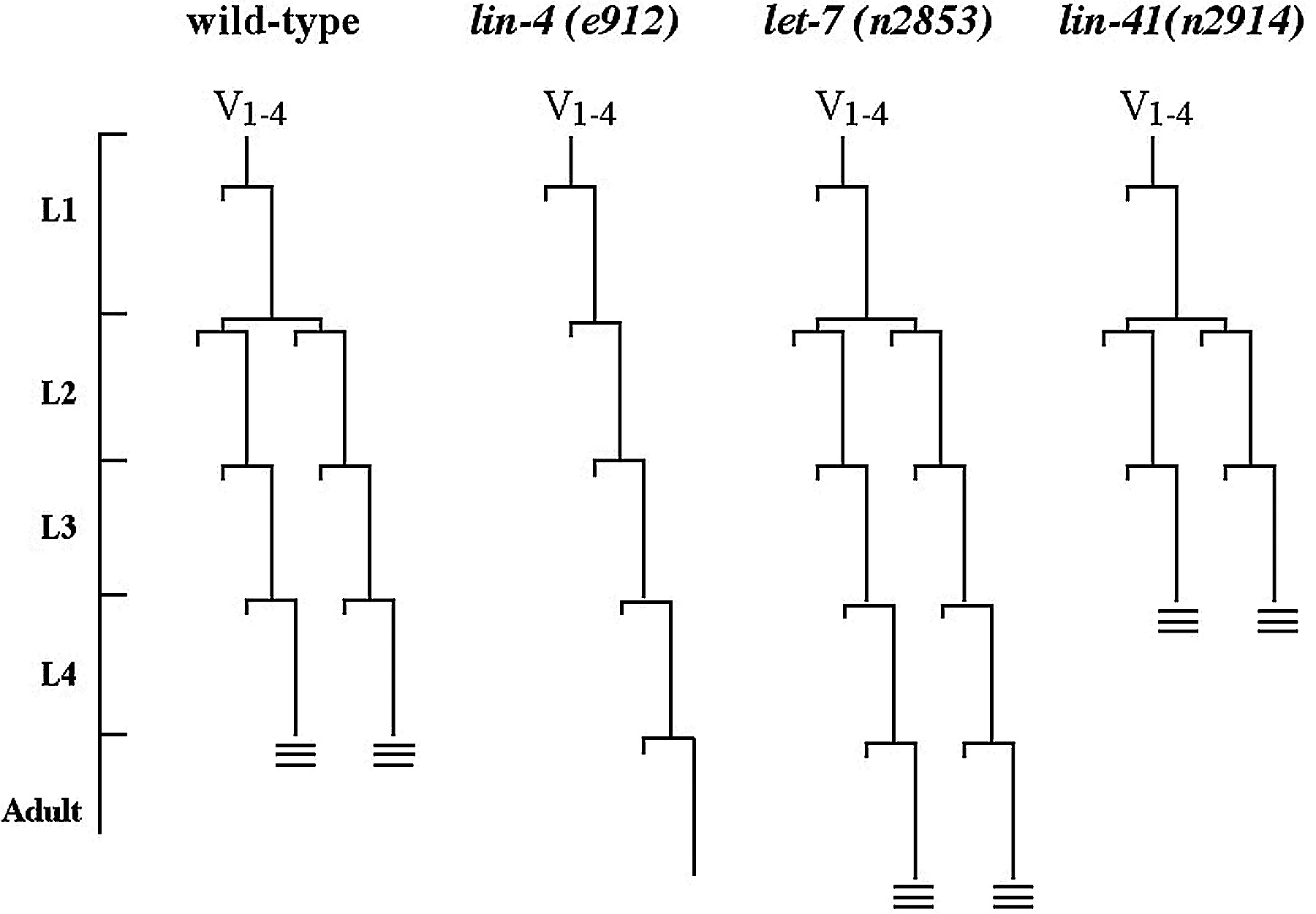
One of lin-4's target genes, lin-14, encodes a novel nuclear protein and is a putative transcription factor (Hong et al., 2000). LIN-14 is present at high levels in newly hatched L1 animals with intense nuclear staining in hypodermal blast cells, which decreases by the L2 stage (Ruvkun and Giusto, 1989). lin-4 loss-of-function (lf) and lin-14 gain-of-function mutants (gf) fail to turn off LIN-14 expression post-L2 and both mutants follow a pattern of retarded development, reiterating the L1 stage patterns. This is due to increased LIN-14 levels (Figure 1) because the lin-4 retarded phenotype depends on the wild-type activity of lin-14 (Ambros, 1989).
lin-14 mRNA levels remain constant throughout development, suggesting post-transcriptional regulation by lin-4 (Wightman et al., 1993). lin-4 is thought to act by antisense base pairing to seven complementary elements in the lin-14 3' UTR to down-regulate lin-14 (Lee et al., 1993; Wightman et al., 1993). All lin-14 mRNA at the L1 and L2 stages is polyribosome associated, suggesting that lin-14 gene regulation does not involve the inaccessibility of lin-14 mRNA to the translational machinery (Olsen and Ambros, 1999). Binding of lin-4 to lin-14 is thought to block protein synthesis or accumulation after the initiation of translation, possibly through a mechanism of targeted LIN-14 degradation (Figure 2; Olsen and Ambros, 1999). However, the precise mechanism by which LIN-14 is down-regulated by lin-4 remains unknown.
The 3' UTR of lin-14 is sufficient for temporal regulation of lin-14, as experiments using a lacZ reporter gene attached to the lin-14 3' UTR, show correct stage specific down-regulation of lacZ at the L1-to-L2 transition (Wightman et al., 1993). lin-14 gf mutations map to the lin-14 3' UTR, and delete the lin-4 complementary elements in the 3' UTR. The lin-4 RNA binds in vitro to a lin-14 3'UTR RNA containing the seven complementary elements but not to a lin-14 mRNA having point mutations in these sites (Ha et al., 1996). Since these mutant sites fail to down-regulate a reporter, these sites are crucial to normal gene regulation. The seven complementary elements in the lin-14 3' UTR are also conserved in C. briggsae, a related Caenorhabditis species (Stein et al., 2003) also suggesting that these sites are important to normal lin-14 regulation. lin-4 also regulates a second gene, lin-28, through a similar mechanism to that for lin-14 (Moss et al., 1997).
The interaction of lin-4 and lin-14 and their effect on the timing of development in C. elegans set a precedent for the study of another miRNA, let-7, which is expressed later in development. let-7 controls the L4-to-adult transition in C. elegans development by interacting with the 3' UTR of multiple target genes, including lin-41. In C. elegans let-7 RNA is expressed at the late L3 and early L4 stages in the hypodermal seam cells (Johnson et al., 2003; Reinhart et al., 2000). let-7 mutants fail to execute the L4-to-adult transition, reiterating larval fates (Figure 1), and die by bursting through the vulva (Reinhart et al., 2000).
The let-7 miRNA is first transcribed as a larger pri-miRNA molecule that is spliced and trans-spliced to SL1 (Bracht et al., 2004). A precursor RNA molecule of about 70 nts is processed into the mature 22 nt molecule by the RNAse III nuclease Dicer (Bernstein et al., 2001; Grishok et al., 2001). Knock down of dcr-1 by RNA interference (RNAi) results in a let-7-like mutant phenotype in which the seam cells fail to terminally differentiate and continue to divide, a pattern characteristic of the retarded hypodermal seam cell development seen in let-7 mutants (Grishok et al., 2001). Grishok et al. also found that correct processing of the lin-4 and let-7 precursors into the shorter functional molecules depends upon dcr-1, with a marked reduction in mature lin-4 and let-7 miRNAs in dcr-1 RNAi animals.
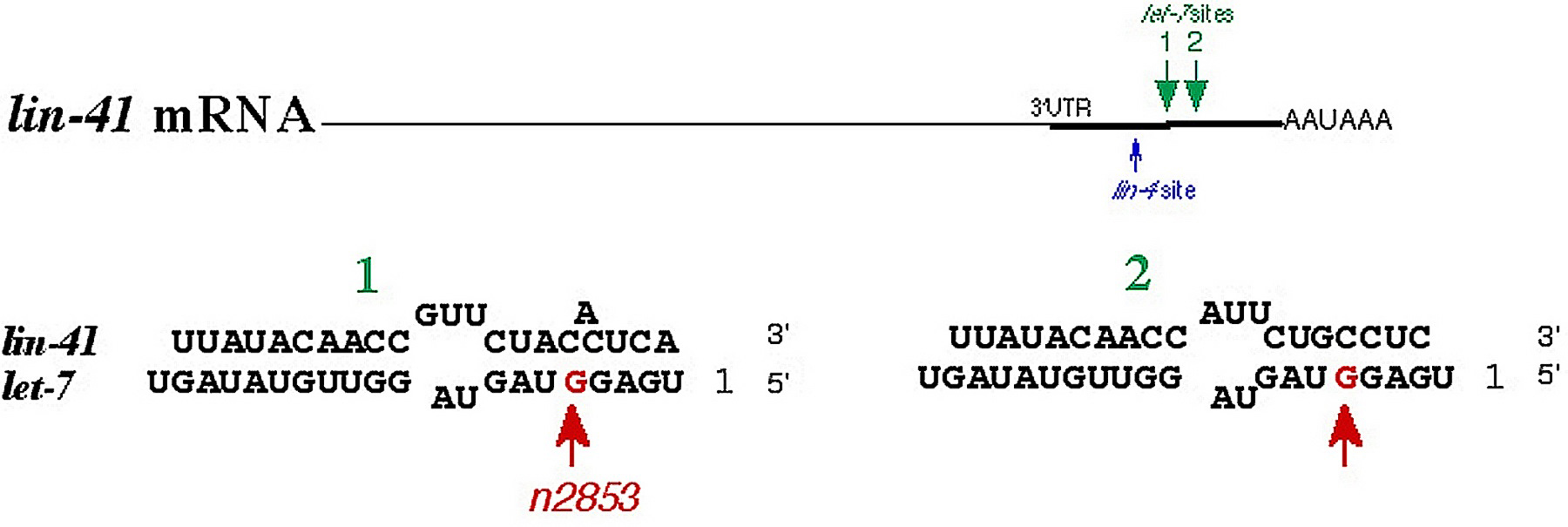
The retarded let-7 phenotype can be partially suppressed by lin-41(lf) mutations, suggesting that lin-41 is repressed by let-7 (Reinhart et al., 2000; Slack et al., 2000). let-7 controls the larval-to-adult transition in C. elegans development by down-regulating LIN-41 and thereby allowing the animals to advance to the adult cell fate programs. LIN-41/GFP is temporally regulated in the hypodermal seam cells, being down-regulated during the L4 stage at the same time as the up-regulation of the let-7 RNA (Slack et al., 2000). The lin-41 3' UTR placed behind a heterologous promoter and a lacZ reporter is sufficient to down-regulate the reporter in a manner similar to that of LIN-41 down-regulation at the L4-to-adult transition (Slack et al., 2000), indicating the importance of the 3'UTR in regulation of lin-41 gene expression. let-7 interacts with two complementary sites in the 3' UTR of lin-41 (Figure 3). Deletion of an 85 base pair region in the lin-41 3' UTR containing the two let-7 complementary sites (LCSs) abrogates this down-regulation, as do let-7 mutations (Reinhart et al., 2000), suggesting that both the LCSs in lin-41 and a wild type copy of let-7 are required for proper down-regulation. A detailed analysis of the let-7/lin-41 interaction reveals that let-7 directly binds to the lin-41 LCSs, and that the context and overall architecture of the sites is important for miRNA function (Vella et al., 2004a; Vella et al., 2004b).
The let-7 sequence is conserved across phylogeny from worms to flies to humans and is expressed in C. elegans, Drosophila and zebrafish, at specific times during development (Pasquinelli et al., 2000). LIN-41 has homologues in flies and vertebrates (Slack et al., 2000) and LCSs are found in Drosophila and zebrafish lin-41 3' UTRs (Pasquinelli et al., 2000). This conservation suggests that this mechanism of gene regulation is also present in larger animals.
The let-7 RNA also exhibits complementarity to the 3' UTR of multiple other genes, including the heterochronic genes lin-14, lin-28, daf-12 and hbl-1, suggesting that let-7 may also bind to these genes and regulate their expression (Grosshans et al., 2005; Lin et al., 2003; Reinhart et al., 2000).
The let-7 miRNA is a member of a family of miRNAs comprised of twelve human let-7 RNAs and four C. elegans RNAs let-7, mir-48, mir-84, and mir-241; Figure 4; Lim et al., 2003a). The RNA sequences of the family members are most highly conserved in the 5' region of the miRNA. It is unclear right now whether these family members have to ability to regulate the same target genes, but the potential appears to exist. One exciting possibility is that the variable 3' region of the miRNA provides some specificity in target interactions. One member of the family, mir-84 is expressed in a dynamic manner in vulval precursor cells and negatively regulates let-60/RAS in these cells (Johnson and Slack, 2005).
While lin-4 and let-7 control temporal patterning, a role for miRNAs in spatial pattern formation has been recently documented. Here, a second case of multiple miRNAs working in a single pathway to govern a biological process in C. elegans elegans has been described. The pathway involves control of left/right asymmetric fates of two neurons in the head (ASE left, ASEL; and ASE right, ASER). lsy-6, is expressed in only one ASE but not the other and regulates the expression of a downstream transcription factor, COG-1, whose 3' UTR contains a partially complementary sequence to the lsy-6 miRNA. The cog-1 3'UTR dictates lsy-6 dependent repression in vivo (Johnston and Hobert, 2003). mir-273, the second miRNA in this pathway, is expressed in the other ASE and exhibits complementarity to the mRNA of the die-1 transcription factor which is regulated post-transcriptionally and expressed mainly in ASEL (Chang et al., 2004). It is interesting to note that the targets of mir-273 and lsy-6 are also regulated at the post-transcriptional level suggesting a common mechanism for miRNAs in regulating C. elegans developmental pathways. It remains to be seen if this is a general feature of C. elegans miRNAs.
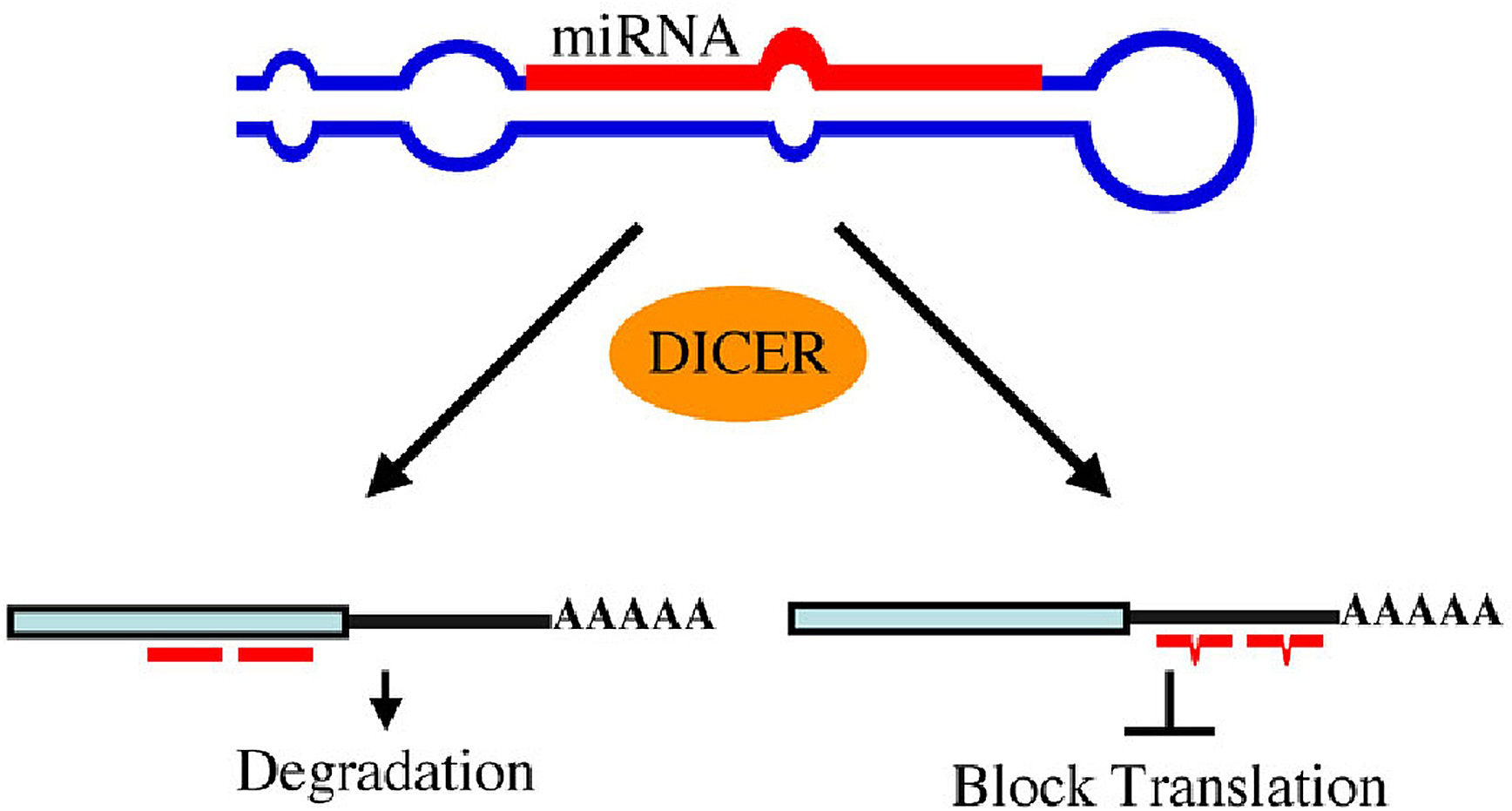
A second class of regulatory RNAs, small interfering RNAs (siRNAs), share several common characteristics with miRNAs. Dicer activity is needed for cleavage of both the longer double-stranded (ds) trigger RNA and the 70-75 nt miRNA precursors that give rise to the mature RNA molecules involved in RNAi (siRNAs) and translational control (miRNAs), respectively. While miRNAs are genomically encoded and siRNAs are derived from exogenous dsRNA, endogenous mRNAs, transposons, viruses, or heterochromatic DNA (Ambros et al., 2003; Bartel, 2004), mature siRNAs and miRNAs are both incorporated into an RNA-induced silencing complex (RISC). The siRNAs act as guides to direct the siRNA::mRNA duplex into the siRNA-induced silencing complex (siRISC; ; Lee et al., 2004b) and the target mRNA is silenced by degradation. The miRNA::mRNA duplex is incorporated into the miRNA-induced silencing complex (miRISC) and the target mRNA is translationally repressed. This may be due to the fact that siRNAs interact with perfect complementarity to their target genes whereas miRNA::target interactions often have bulges or several mismatched bases.
RISC contains members of the Argonaute (Ago) family of proteins (Hammond et al., 2001). Other RISC-associated proteins include VIG and Fragile X-related protein (Caudy et al., 2002), two RNA-binding proteins and Tudor-SN, a nuclease (Caudy et al., 2003). Members of the Ago family co-immunoprecipate with miRNAs (Carmell et al., 2002; Caudy et al., 2002; Mourelatos et al., 2002). In particular, ALG-1 and ALG-2 appear to function with lin-4 and let-7 to control developmental timing (Grishok et al., 2001). One idea is that there are many RISC complexes, all of which contain common core components, but have specificity proteins, such as Ago proteins (which are very numerous in C. elegans with about 24 (Bartel, 2004; Carmell et al., 2002)that distinguish siRISC/miRISC complexes and their consequent modes of regulation.
|
Figure 5. miRNAs use two mechanisms to exert gene regulation. Some animal miRNAs can bind to mRNA targets with exact complementarity and induce the RNAi pathway. miRNAs also bind to targets with imperfect complementarity and block translation. There is no evidence that C. elegans miRNAs use the former pathway.
MiRNAs were discovered in C. elegans. Additionally, much of what we know about miRNAs and their targets has come from studies in C. elegans . Even though C. elegans has over 100 miRNAs, the functions of only five have been elucidated by genetic studies, and fewer than fifteen valid C. elegans target genes are known. Much still remains to be discovered and many unanswered questions remain. For example, what is the role of the other C. elegans miRNAs, what are their targets, how do they function and do these miRNAs work similarly in higher organisms? It seems clear from current studies that miRNAs play important roles in C. elegans development; while studies in other organisms show that they are important regulators of metabolism. It will not be surprising to learn that most biological processes are touched in some way by miRNAs. The excitement generated by the discovery of miRNAs, and their similarity to siRNAs, has made a huge impact on our thinking about genetic control. Certainly, the geneticist mapping interesting mutations has to now be aware of non-protein coding genes mapping in their genetic intervals, and the molecular biologist has to consider the possibility that their favorite gene is regulated by a miRNA. miRNAs may prove to be only a small part of a larger emerging picture of RNA-based gene regulation (Ambros, 2003; Winkler et al., 2002).
Ambros, V. (1989). A hierarchy of regulatory genes controls a larva-to-adult developmental switch in C. elegans . Cell 57, 49–57. Abstract Article
Ambros, V. (2003). MicroRNA pathways in flies and worms: growth, death, fat, stress, and timing. Cell 113, 673–676. Abstract Article
Ambros, V., and Horvitz, H.R. (1984). Heterochronic mutants of the nematode Caenorhabditis elegans. Science 226, 409–416. Abstract
Ambros, V., Lee, R.C., Lavanway, A., Williams, P.T., and Jewell, D. (2003). MicroRNAs and other tiny endogenous RNAs in C. elegans . Curr. Biol. 13, 807–818. Article
Bartel, D.P. (2004). MicroRNAs: genomics, biogenesis, mechanism, and function. Cell 116, 281–297. Abstract Article
Bernstein, E., Caudy, A.A., Hammond, S.M., and Hannon, G.J. (2001). Role for bidentate ribonuclease in the initiation step of RNA interference. Nature 409, 295–296. Article
Bracht, J., Hunter, S., Eachus, R., Weeks, P., and Pasquinelli, A.E. (2004). Trans-splicing and polyadenylation of let-7 microRNA primary transcripts. Rna 10, 1586–1594. Abstract Article
Carmell, M.A., Xuan, Z., Zhang, M.Q., and Hannon, G.J. (2002). The Argonaute family: tentacles that reach into RNAi, developmental control, stem cell maintenance, and tumorigenesis. Genes Dev. 16, 2733–2742. Abstract Article
Caudy, A.A., Ketting, R.F., Hammond, S.M., Denli, A.M., Bathoorn, A.M., Tops, B.B., Silva, J.M., Myers, M.M., Hannon, G.J., and Plasterk, R.H. (2003). A micrococcal nuclease homologue in RNAi effector complexes. Nature 425, 411–414. Abstract Article
Caudy, A.A., Myers, M., Hannon, G.J., and Hammond, S.M. (2002). Fragile X-related protein and VIG associate with the RNA interference machinery. Genes Dev. 16, 2491–2496. Abstract Article
Chalfie, M., Horvitz, H.R., and Sulston, J.E. (1981). Mutations that lead to reiterations in the cell lineages of C. elegans . Cell 24, 59–69. Article
Chang, S., Johnston, R.J., Jr., Frokjaer-Jensen, C., Lockery, S., and Hobert, O. (2004). MicroRNAs act sequentially and asymmetrically to control chemosensory laterality in the nematode. Nature 430, 785–789. Abstract Article
Feinbaum, R., and Ambros, V. (1999). The timing of lin-4 RNA accumulation controls the timing of postembryonic developmental events in Caenorhabditis elegans. Dev. Biol. 210, 87–95. Abstract Article
Grad, Y., Aach, J., Hayes, G.D., Reinhart, B.J., Church, G.M., Ruvkun, G., and Kim, J. (2003). Computational and experimental identification of C. elegans microRNAs. Mol. Cell 11, 1253–1263. Abstract Article
Grishok, A., Pasquinelli, A.E., Conte, D., Li, N., Parrish, S., Ha, I., Baillie, D.L., Fire, A., Ruvkun, G., and Mello, C.C. (2001). Genes and mechanisms related to RNA interference regulate expression of the small temporal RNAs that control C. elegans developmental timing. Cell 106, 23–34. Abstract Article
Grosshans, H., Johnson, T., Reinert, K.L., Gerstein, M., and Slack, F.J. (2005). The temporal patterning microRNA let-7 regulates several transcription factors at the larval to adult transition in C. elegans. Dev. Cell 8, 321–330. Abstract Article
Ha, I., Wightman, B., and Ruvkun, G. (1996). A bulged lin-4/lin-14 RNA duplex is sufficient for Caenorhabditis elegans lin-14 temporal gradient formation. Genes Dev. 10, 3041–3050.
Hammond, S.M., Boettcher, S., Caudy, A.A., Kobayashi, R., and Hannon, G.J. (2001). Argonaute2, a link between genetic and biochemical analyses of RNAi. Science 293, 1146–1150. Abstract Article
Hong, Y., Lee, R.C., and Ambros, V. (2000). Structure and function analysis of LIN-14, a temporal regulator of postembryonic developmental events in Caenorhabditis elegans. Mol. Cell Biol. 20, 2285–2295. Abstract Article
Houbaviy, H.B., Murray, M.F., and Sharp, P.A. (2003). Embryonic stem cell-specific MicroRNAs. Dev. Cell 5, 351–358. Abstract Article
Johnson, S.M., Lin, S.Y., and Slack, F.J. (2003). The time of appearance of the C. elegans let-7 microRNA is transcriptionally controlled utilizing a temporal regulatory element in its promoter. Dev. Biol. 259, 364–379. Abstract Article
Johnson, S.M., and Slack, F.J. (2005). RAS is regulated by the let-7 miRNA family. Cell 120, 635–647. Article
Johnston, R.J., and Hobert, O. (2003). A microRNA controlling left/right neuronal asymmetry in Caenorhabditis elegans. Nature 426, 845–849. Abstract Article
Lagos-Quintana, M., Rauhut, R., Lendeckel, W., and Tuschl, T. (2001). Identification of novel genes coding for small expressed RNAs. Science 294, 853–858 Article
Lagos-Quintana, M., Rauhut, R., Yalcin, A., Meyer, J., Lendeckel, W., and Tuschl, T. (2002). Identification of tissue-specific microRNAs from mouse. Curr. Biol. 12, 735–739. Abstract Article
Lau, N.C., Lim, L.P., Weinstein, E.G., and Bartel, D.P. (2001). An abundant class of tiny RNAs with probable regulatory roles in Caenorhabditis elegans. Science 294, 858–862. Abstract Article
Lee, R.C., and Ambros, V. (2001). An extensive class of small RNAs in Caenorhabditis elegans. Science 294, 862–864. Article
Lee, R.C., Feinbaum, R.L., and Ambros, V. (1993). The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 75, 843–854. Abstract Article
Lee, Y., Ahn, C., Han, J., Choi, H., Kim, J., Yim, J., Lee, J., Provost, P., Radmark, O., Kim, S., and Kim, V.N. (2003). The nuclear RNase III Drosha initiates microRNA processing. Nature 425, 415–419. Abstract Article
Lee, Y., Jeon, K., Lee, J.T., Kim, S., and Kim, V.N. (2002). MicroRNA maturation: stepwise processing and subcellular localization. EMBO J. 21, 4663–4670. Abstract Article
Lee, Y., Kim, M., Han, J., Yeom, K.H., Lee, S., Baek, S.H., and Kim, V.N. (2004a). MicroRNA genes are transcribed by RNsA polymerase II. EMBO J. Abstract Article
Lee, Y.S., Nakahara, K., Pham, J.W., Kim, K., He, Z., Sontheimer, E.J., and Carthew, R.W. (2004b). Distinct roles for Drosophila Dicer-1 and Dicer-2 in the siRNA/miRNA silencing pathways. Cell 117, 69–81. Abstract Article
Lim, L.P., Lau, N.C., Weinstein, E.G., Abdelhakim, A., Yekta, S., Rhoades, M.W., Burge, C.B., and Bartel, D.P. (2003). The microRNAs of Caenorhabditis elegans. Genes Dev. 17, 991–1008. Abstract Article
Lin, S.Y., Johnson, S.M., Abraham, M., Vella, M.C., Pasquinelli, A., Gamberi, C., Gottlieb, E., and Slack, F.J. (2003). The C elegans hunchback homolog, hbl-1, controls temporal patterning and is a probable microRNA target. Dev. Cell 4, 639–650. Abstract Article
Moss, E.G., Lee, R.C., and Ambros, V. (1997). The cold shock domain protein LIN-28 controls developmental timing in C. elegans and is regulated by the lin-4 RNA. Cell 88, 637–646. Abstract Article
Mourelatos, Z., Dostie, J., Paushkin, S., Sharma, A., Charroux, B., Abel, L., Rappsilber, J., Mann, M., and Dreyfuss, G. (2002). miRNPs: a novel class of ribonucleoproteins containing numerous microRNAs. Genes Dev. 16, 720–728. Abstract Article
Olsen, P.H., and Ambros, V. (1999). The lin-4 regulatory RNA controls developmental timing in Caenorhabditis elegans by blocking LIN-14 protein synthesis after the initiation of translation. Dev. Biol. 216, 671–680. Abstract Article
Pasquinelli, A.E., Reinhart, B.J., Slack, F., Martindale, M.Q., Kuroda, M.I., Maller, B., Hayward, D.C., Ball, E.E., Degnan, B., Muller, P., et al. (2000). Conservation of the sequence and temporal expression of let-7 heterochronic regulatory RNA. Nature 408, 86–89. Abstract Article
Reinhart, B.J., Slack, F.J., Basson, M., Pasquinelli, A.E., Bettinger, J.C., Rougvie, A.E., Horvitz, H.R., and Ruvkun, G. (2000). The 21-nucleotide let-7 RNA regulates developmental timing in Caenorhabditis elegans. Nature 403, 901–906. Abstract Article
Reinhart, B.J., Weinstein, E.G., Rhoades, M.W., Bartel, B., and Bartel, D.P. (2002). MicroRNAs in plants. Genes Dev. 16, 1616–1626. Abstract Article
Ruvkun, G., and Giusto, J. (1989). The Caenorhabditis elegans heterochronic gene lin-14 encodes a nuclear protein that forms a temporal developmental switch. Nature 338, 313–319. Abstract Article
Slack, F.J., Basson, M., Liu, Z., Ambros, V., Horvitz, H.R., and Ruvkun, G. (2000). The lin-41 RBCC gene acts in the C. elegans heterochronic pathway between the let-7 regulatory RNA and the LIN-29 transcription factor. Mol. Cell 5, 659–669. Abstract Article
Stein, L.D., Bao, Z., Blasiar, D., Blumenthal, T., Brent, M.R., Chen, N., Chinwalla, A., Clarke, L., Clee, C., Coghlan, A., et al. (2003). The Genome Sequence of Caenorhabditis briggsae: A Platform for Comparative Genomics. PLoS. Biol. 1, E45. Abstract Article
Vella, M.C., Choi, E.Y., Lin, S.Y., Reinert, K., and Slack, F.J. (2004a). The C. elegans microRNA let-7 binds to imperfect let-7 complementary sites from the lin-41 3'UTR. Genes Dev. 18, 132–137. Abstract Article
Vella, M.C., Reinert, K., and Slack, F.J. (2004b). Architecture of a validated microRNA:target interaction. Chem. Biol. 11, 1619–1623 Abstract Article
Wickens, M., and Takayama, K. (1994). RNA. Deviants--or emissaries. Nature 367, 17–18. Abstract Article
Wightman, B. Ha, I., and Ruvkun, G. (1993). Posttranscriptional regulation of the heterochronic gene lin-14 by lin-4 mediates temporal pattern formation in C. elegans . Cell 75, 855–862. Abstract Article
*Edited by Thomas Blumenthal. Last revised March 16, 2005. Published September 21, 2005. This chapter should be cited as: Vella, M.C. and Slack, F.J. C. elegans microRNAs (September 21, 2005), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.26.1, http://www.wormbook.org.
Copyright: © 2005 Monica C. Vella and Frank J. Slack. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
§To whom correspondence should be addressed. E-mail: frank.slack@yale.edu
 All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.
All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.