Embryo series courtesy of Einhard Schierenberg
Embryo series courtesy of Einhard SchierenbergAbstract
Four biogenic amines: octopamine, tyramine, dopamine and serotonin act in C. elegans to modulate behavior in response to changing environmental cues. These neurotransmitters act at both neurons and muscles to affect egg laying, pharyngeal pumping, locomotion and learning. A variety of experimental approaches including genetic, imaging, biochemical and pharmacological analyses have been used to identify the enzymes and cells that make and release the amines and the cells and receptors that bind them. Dopamine and serotonin act through receptors and downstream signaling mechanisms similar to those that operate in the mammalian brain suggesting that C. elegans will provide a valuable model for understanding biogenic amine signaling in the brain.
The biogenic amines dopamine, serotonin, octopamine and tyramine can all be detected in C. elegans extracts by HPLC analysis and appear to function as neurotransmitters or neuromodulators (Sulston et al., 1975; Sanyal et al., 2004; Horvitz et al., 1982; Alkema et al., 2005). Other signaling amines found in vertebrates (histamine, epinephrine, and norepinephrine) appear not to be made in C. elegans (Horvitz et al., 1982; Sanyal et al., 2004). Biogenic amines modulate a variety of C. elegans behaviors including locomotion, egg laying (see Egg laying), defecation, and foraging. Dopamine and serotonin have established roles in mammalian nervous system function and abnormal signaling by these neurotransmitters has been implicated in several human diseases including Parkinson's disease, schizophrenia, and depression. It is not clear whether octopamine or its biosynthetic precursor tyramine function as neurotransmitters in mammals. Octopamine, however, can act as a neurotransmitter in invertebrates and recent evidence has implicated tyramine as a neurotransmitter in C. elegans (Alkema et al., 2005). Studies in C. elegans have begun to elucidate the molecular and cellular signaling mechanisms by which these neurotransmitters control behavior.
A number of tools are available to study the molecular and cellular mechanisms of signaling by biogenic amines in C. elegans. The genes encoding the enzymes responsible for synthesis of these neurotransmitters and the gene encoding the transporter (cat-1) that loads these neurotransmitters into vesicles have been identified and mutants are available in each (see Figure 1). Analysis of the phenotypes of such mutants has shed light on the behaviors controlled by the amines and detailed analysis of the expression patterns of the biosynthetic enzymes and transporter has helped identify the cells and neurons in which the neurotransmitters are made and released (see Table 1). Cells containing dopamine and serotonin can also be identified using formaldehyde-induced fluorescence and / or antibody stains. Seventeen putative biogenic amine receptors have been identified based on sequence similarity to mammalian receptors, and mutants for most of these are available (see Table 2). Although the specific biogenic amine that activates a given receptor homolog cannot be accurately predicted based purely on sequence analysis, in a growing number of cases pharmacological analysis of cloned C. elegans receptors has been used to determine their likely physiological ligands (see Table 2). Transgenes in which the promoters for the receptors drive expression of fluorescent proteins have been used to identify the cells in which the receptors are expressed and that thus respond to the neurotransmitter signal (see Table 2). Finally, a variety of drugs that target biogenic amine receptors are available and their effects on behavior can be helpful in determining the physiological roles of the receptors (see Table 3). The specificity of these drugs on C. elegans receptors is however largely unclear, as most of the drugs were developed against mammalian receptors and interact with regions of the receptors that may not be evolutionarily conserved. As a result, many of the drugs tested show different affinities for C. elegans receptors than seen for their mammalian receptor counterparts, and indeed may have completely different activities at the C. elegans receptors than they have at the mammalian receptors.
This chapter provides a reference source that organizes the growing body of literature in the field of biogenic amine signaling in C. elegans and also highlights current gaps in the field.
|
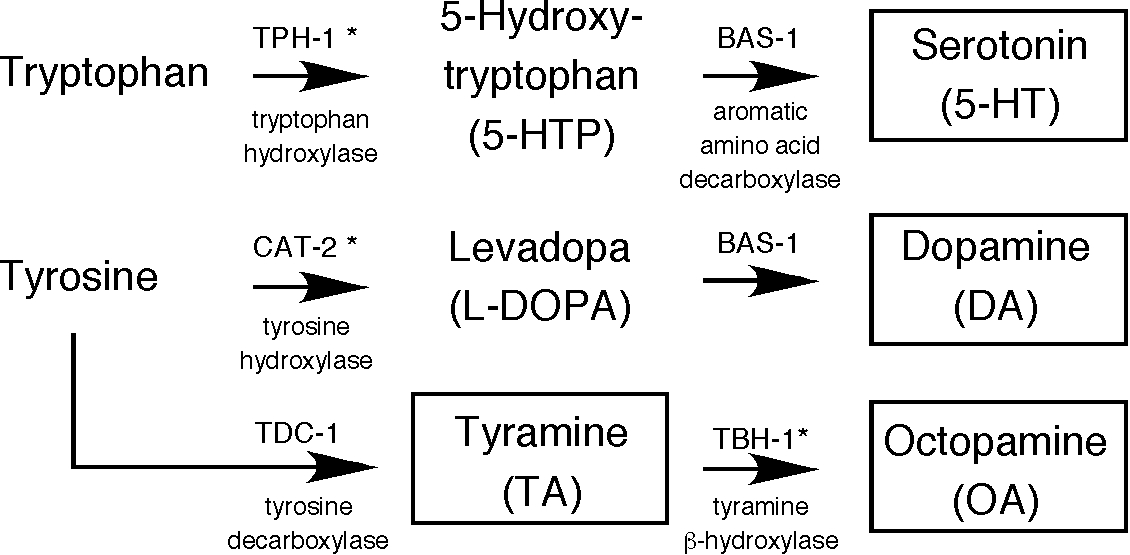
Figure 1. Biosynthetic pathways for biogenic amines. Asterisks indcate enzymes that require a cofactor whose synthesis requires a GTP cyclohydrolase I encoded by the cat-4 gene (Sulston et al., 1975; Loer and Kenyon, 1993). cat-4 mutants have reduced formaldehyde induced fluroscence staining for dopamine and serotonin. References are as follows: TPH-1: Sze et al., 2000; BAS-1: Loer and Kenyon, 1993; CAT-2: Lints and Emmons, 1999; TDC-1: Alkema et al., 2005; TBH-1: Alkema et al., 2005.
Table 1. Cells that synthesize biogenic amines
| Neuron / cell | Dopamine | Serotonin | Octopamine | Tyramine | CAT-1 expressiona |
|---|---|---|---|---|---|
| ADE | +1, 9, 10, 11 | − | − | − | + |
| PDE | +1, 9, 10, 11 | − | − | − | + |
| CEP | +1, 9, 10, 11 | − | − | − | + |
| R5A, R7A, R9A | +2, 11 | − | − | − | NE |
| RIC | − | − | +3 | +3 | + |
| Gonad sheath | − | − | +3 | +3 | NR |
| RIM | − | − | − | +3 | NR |
| UV1 | − | − | − | +3 | NR |
| CP1-6 | − | +4,12 | − | − | NE |
| R1, R3, R9 | − | +4 | − | − | NE |
| NSM | − | +5, 6, 8, 12 | − | − | + |
| HSN | − | +6, 8, 12 | − | − | + |
| VC4, 5 | − | +7 | − | − | + |
| ADF | − | +7, 12 | − | − | + |
| RIH | − | +7, 12 | − | − | + |
| AIM | − | +7, 12 | − | − | + |
| CAN | − | − | − | − | + |
| aAll CAT-1 expression was reported in a single publication (Duerr et al., 1999) NE, not examined; NR, not reported. | |||||
References
| 1. Sulston et al., 1975 |
| 2. Sulston and Horvitz, 1977 |
| 3. Alkema et al., 2005 |
| 4. Loer and Kenyon, 1993 |
| 5. Horvitz et al., 1982 |
| 6. Desai et al., 1988 |
| 7. Duerr et al., 1999 |
| 8. McIntire et al., 1992 |
| 9. Lynch et al., 1995 |
| 10. Nass et al., 2002 |
| 11. Lints and Emmons 1999 |
| 12. Sze et al., 2000 |
Octopamine is synthesized from tyramine by the tyramine β-hydroxylase TBH-1. Based on the expression of TBH-1 as determined by antibody stain, octopamine synthesis is limited to the RIC interneurons and the gonadal sheath cells, (Alkema et al., 2005). The behavioral defects of tbh-1 mutants have not been described in detail, however they share several (though apparently not all) pleiotropies with tdc-1 mutants (which cannot synthesize tyramine or octopamine, see Figure 1). One of the issues in this field is whether the cells that make octopamine also release tyramine as a neurotransmitter, or whether tyramine functions in those cells simply as a precursor in octopamine synthesis. While no octopamine receptors have yet been verified using pharmacological criteria, SER-3 appears to be a leading candidate and may be activated by octopamine in the SIA interneurons in response to starvation (Suo et al., 2006). Behavioral defects associated with ser-3 mutants have not yet been described, however treatment of animals with exogenous octopamine causes several behavioral effects, including inhibition of egg laying and inhibition of pharyngeal pumping (Horvitz et al., 1982; Alkema et al., 2005). Both of these behaviors are normally stimulated by the presence of food, suggesting that octopamine signaling opposes the signal induced by food. Supporting the role of octopamine in the control of egg laying, treatment with phentolamine (an octopamine antagonist in insects) stimulates egg-laying behavior (Horvitz et al., 1982).
Tyramine is present in low abundance in C. elegans, perhaps because it is made in only a few cells, and in some of those cells serves as an intermediate in the synthesis of octopamine (Alkema et al., 2005). The enzyme used to synthesize tyramine (TDC-1) appears to be expressed in only two cells types that do not also express the enzyme that converts tyramine to octopamine (TBH-1), and these cells (the RIM-1 motorneurons and the uv1 neuroendocrine cells) may thus release tyramine as a neurotransmitter (Alkema et al., 2005). tdc-1 mutants exhibit behavioral defects not shared with tbh-1 mutants, and characterization of these defects indicate that tyramine is required for the following: 1) inhibition of head oscillations as animals back in response to light touch to the anterior portion of the body; 2) inhibition of egg laying; and 3) modulation of spontaneous reversals (Alkema et al., 2005; Rex et al., 2004). The latter two functions suggest that tyramine may be preferentially released during times of low food abundance when the animal needs to reduce egg laying and search for new food supplies. Two receptors, SER-2 and TYRA-2, have been identified that bind tyramine with relatively high affinity when expressed in cell culture (Rex et al., 2002; Rex et al., 2004; Rex et al., 2005). Mutations in ser-2 cause several behavioral defects (see Table 2), some of which are similar to the behavioral defects seen in tdc-1 mutants. For example, ser-2 mutants fail to suppress head oscillations in response to touch (Rex et al., 2004). Signaling to suppress head oscillations acts through the mechanosensory neurons ALM and AVM, which form gap junctions with the command interneurons AVA and AVD that control backward movement. The AVA and AVD neurons in turn form gap junctions with the tyraminergic RIM neurons which then synapse onto and presumably modulate the activity of the RMD motorneurons that directly innervate head muscles (Alkema et al., 2005). Interestingly, none of the cells that have been identified that express SER-2 are postsynaptic to the RIM motorneurons, suggesting that if tyramine acts through SER-2 to suppress head oscillations it must function extrasynaptically. SER-2 is expressed on the head muscles that effect head movements, and thus these muscles are likely sites of action for tyramine (Tsalik et al., 2003). While a single tyra-2 mutant is available, an analysis of TYRA-2 effects on behavior has not yet been published. TYRA-2 is expressed in neurons of the amphid sensilla (ASE, ASG, ASH and ASI), the ALM mechanosensory neurons and a few other neurons (Rex et al., 2005). The expression of TYRA-2 on the ALM neurons suggests that tyramine could inhibit head oscillations by modulating the activity of the ALM neurons, which in turn synapse onto and presumably modulate the activity of the RMD neurons. The ALM neurons are not postsynaptic to the neurons that make tyramine and thus tyramine would have to act extrasynaptically to act through ALM-expressed TYRA-2 receptors.
Exogenous tyramine inhibits serotonin-stimulated pharyngeal pumping, and mutations in ser-2 block this effect, consistent with the idea that tyramine is released in response to food deprivation to inhibit pharyngeal pumping. SER-2 is expressed on pharyngeal muscles and thus tyramine could act directly on them (Tsalik et al., 2003). Exogenous tyramine also inhibits egg laying in wild-type animals, and tdc-1 mutants are weakly hyperactive egg layers (Alkema et al., 2005). If tyramine does inhibit egg laying it likely does not act through SER-2, as tyramine still inhibits egg laying in ser-2 mutants. While the synthesis of tyramine in the gonadal sheath cells is consistent with a role in egg laying, tyramine may simply be an intermediate in those cells in the synthesis of octopamine. Indeed, the inhibitory effects of exogenous tyramine on egg-laying behavior may be the result of tyramine being converted to octopamine inside the animal, as the effects of exogenous tyramine have not been tested in tbh-1 mutants, and exogenous octopamine also inhibits egg laying.
Table 2. Biogenic amine receptors: their expression patterns, pharmacology and mutant phenotypes
| Clone / Gene namea | Expression patternsb | Pharmacology | Mutants | Mutant phenotype |
|---|---|---|---|---|
| F15A8.5 (dop-1) | ALM, ALN, AVM, PLN, PVQ, RIS and in other head neurons, head muscles, excretory gland cells, amphid and labial support cells7. Cholinergic ventral cord motor neurons9. ALM, PHC, PLM, and unidentified others11. | DA > 5-HT > OA > TA4. | ev748, ok398, tm534, tm535, vs100, vs101 | Defective in tap habituation11. Defective in basal slowing and response to exogenous dopamine (in dop-3 background)9. |
| K09G1.4 (dop-2) | ADE, CEP, PDE, and other head and tail neurons3. PDA, RIA and other head neurons, RID, SIA, SIB7. | DA > 5-HT > TA > OA3. | tm1062, vs105 | ND |
| T14E8.3 (dop-3) | Cholinergic and GABA’ergic ventral cord motor neurons9. | ND | vs106, tm1356 | Defective in basal slowing and response to exogenous dopamine9. |
| F59C12.2 (ser-1) | pm3, pm4, pm5, pm6, pm7, pm87. PVT, pharyngeal muscles, vulval muscles and other head, tail and some ray neurons10. Diagonal muscles of male, pharyngeal muscles, neurons of the head, neurons in the tail including PVT and PVQ, ventral cord motor neurons (not VCs), vulval muscles, uterine cells and cells of the posterior intestine21. Neurons in the head20. | 5-HT > DA, OA2. | ok345 | Defective in egg laying in response to 5-HT10, 20, 21 and fluoxetine10. Males defective in tail curling in response to 5-HT21. |
| Y22D7AR.13 (ser-4) | PVT, RIB, RIS and other head neurons, a pharyngeal neuron, and a tail neuron7. Neurons in the head20. | 5-HT > DA, OA1. | ok512 | Defective in egg laying in response to imipramine10. Males defective in tail curling in response to 5-HT20. |
| C09B7.1 (ser-7) | M4 motor neuron5. Pharyngeal neurons including MC, M4, I2, I3, I4, I6, M2, M3, M5, vulval muscles19. | 5-HT5; 5-HT > DA, TA, OA19. | tm1325, tm1548, tm1728, ok1944 | Defective in egg laying in response to 5-HT19. |
| K06C4.6 (mod-1) | Head, ventral cord and tail neurons, not in muscle cells12. | 5-HT > > DA, TA, OA12. | ok103 | Defective in enhanced slowing12, 13. |
| C02D4.2 (ser-2) | ser-2A: AIY, ALN, AUA, AVH, CAN, DA9, LUA, NSM, PVC, RIA, RIC, RID, SABD, SABV, SDQ, head muscles, diagonal muscles, pm1/6 muscles7, ser-2b: AIY, AIZ, BDU, DVA, PVT, RID, RME, SIAD, SIAV, excretory gland cells7. ser-2d: OLL, PVD7. ser-2(see footnote (c)):RME, RID, BDU, AIY, AVH, AIZ, ALN, RIC, RIA, PDA, ut1 and ut28. | ser-2c TA > OA = DA > 5-HT6, 8; ser-2Ac TA > OA > 5-HT > DA8. | pk1357 | Defective in egg laying in response to 5-HT10. Fail to suppress head oscillations while reversing8. Blocks 5-HT stimulated pharyngeal pumping8. |
| K02F2.6 (ser-3) | Neurons of the head including SIA and the tail neurons PHA, PHB, PVQ, head muscles, intestine, phasmid socket cells, spermatheca, eggs, gonad and vulva17. | ND | ad1774 | ND |
| T02E9.3 | ND | ND | ok568 | ND |
| C52B11.3 (dop-4) | Pharyngeal neurons I1 and I2, ASG, AVL, CAN, PQR, male ray 8, cells of the vulva, intestine, rectal gland and rectal epithelium16. | ND | ok1321, tm1392 | ND |
| M03F4.3 | Some neurons in the head and in the vulva20. | ND | ok325 | ND |
| F14D12.6 | ND | ND | ok371 | ND |
| F01E11.5 (tyra-2) | Pharyngeal neurons MC and NSM, amphid neurons ASE, ASG, ASH, ASI, and other neurons including PVD, CAN, and ALM. | TA > OA = DA > 5-HT18 | tm1815, tm1846 | ND |
| Y54G2A.35 | ND | ND | tm2146, tm2104 | ND |
| C24A8.1 | ND | ND | - | ND |
| F16D3.7 | Body-wall muscles, vulval muscles and some neurons in the head20. | ND | tm2647 | ND |
| aBiogenic amine receptors identified by blast searches of the C. elegans genomic and protein databases14, 15, 9, 7. |
| bExpression patterns of biogenic amine receptors have been inferred from analysis of transgenes in which the promoter region for each receptor gene controlled the expression of fluorescent proteins. In certain cases the expression pattern of a particular receptor was examined by different investigators using different promoter regions. This resulted in different expression patterns. In such cases we have included the expression pattern observed by each investigator separately followed by the publication reference. |
| cThe ser-2 and ser-2A isoforms described by Rex et al. are different from the ser-2 and ser-2A isoforms described by Tsalik et al. |
References
| 1. Olde and McCombie, 1997 |
| 2. Hamdan et al., 1999 |
| 3. Suo et al., 2003 |
| 4. Suo et al., 2002 |
| 5. Hobson et al., 2003 |
| 6. Rex and Komuniecki, 2002 |
| 7. Tsalik et al., 2003 |
| 8. Rex et al. 2004 |
| 9. Chase et al., 2004 |
| 10. Dempsey et al., 2005 |
| 11. Sanyal et al., 2004 |
| 12. Ranganathan et al., 2000 |
| 13. Sawin et al., 2000 |
| 14. Bargmann, 1998 |
| 15. Komuniecki et al., 2004 |
| 16. Sugiura et al., 2005 |
| 17. Suo et al., 2006 |
| 18. Rex et al., 2005 |
| 19. Hobson et al., 2006 |
| 20. Carre-Pierrat et al., 2006 |
| 21. Carnell et al., 2006 |
Dopamine is made in eight neurons in the hermaphrodite and in an additional six neurons located in the tail of the male (see Table 1). Each of these fourteen neurons are thought to be mechanosensory, and ablation of these cells or mutations that block synthesis or release of dopamine cause defects in the animal's ability to sense or respond to changes in its environment (see Mechanosensation; Sawin et al., 2000; Hills et al., 2004; Sanyal et al., 2004; Liu and Sternberg, 1995; Loer and Kenyon, 1993; Bettinger and McIntire, 2004; Duerr et al., 1999). Dopamine signaling has established roles in the modulation of locomotion behavior and in learning. Dopamine-dependent effects on C. elegans behavior have been gleaned primarily from the analysis of: 1) wild-type animals exposed to exogenous dopamine; 2) wild-type animals in which the dopaminergic neurons have been ablated; and 3) mutants for cat-2, which encodes a tyrosine hydroxylase responsible for catalyzing the rate-limiting step in the synthesis of dopamine (see Figure 1; Schafer and Kenyon, 1995; Weinshenker and Thomas, 1995; Sawin et al., 2000; Hills et al., 2004; Liu and Sternberg, 1995; Sanyal et al., 2004; Loer and Kenyon, 1993; Bettinger and McIntire, 2004).
Dopamine signaling allows C. elegans to respond to changes in its environment by modulating locomotion behavior. Well-fed, wild-type animals slow their locomotion rate when they encounter a bacterial lawn and this “basal slowing response” requires dopamine, as cat-2 mutants or animals in which the dopaminergic neurons have been ablated fail to exhibit basal slowing (Sawin et al., 2000). Consistent with the idea that dopaminergic neurons are mechanosensory, this slowing response appears to be caused by physical rather than chemical attributes of the bacterial lawn as wild-type animals also slow in response to a Sephadex matrix and this slowing response also requires cat-2 (Sawin et al., 2000).
Dopamine signaling also allows animals to search efficiently for new food sources. Well-fed, wild-type animals after exhausting a food source search the immediate area for additional food before expanding their search to include more distant, surrounding areas. This localized search for food is called area restricted searching and is accomplished in C. elegans through an increased frequency of high-angled turns immediately after an animal has exhausted a food supply. If, after a short time, the animal fails to find additional food in the area where it had been feeding, the frequency of high-angled turns is reduced and, as a result, the animal begins to explore more distant areas. Dopamine signaling is required for area restricted searching as cat-2 mutants or animals in which the dopaminergic neurons have been ablated do not increase the frequency of high-angled turns immediately after exhausting their food supply like wild-type animals and thus spend less time in the immediate area in which food was recently exhausted. Consistent with this role for dopamine signaling, animals exposed to exogenous dopamine display an increased frequency of high-angled turns and animals exposed to raclopride, a dopamine antagonist, display a decreased frequency of high-angled turns (Hills et al., 2004).
Dopamine thus plays at least two distinct roles in the modulation of locomotion behavior to ensure that animals remain in the area of food. This ability to stay near food is vital for C. elegans to survive in the wild, and animals that briefly wander off a food source must quickly relocate it. In such animals dopamine signals to cause an increased frequency of high-angled turns that prevent the animal from wandering too far from the food source. Upon re-encountering the food source dopamine signals to cause a slowing of forward locomotion increasing the likelihood that the animal does not wander away from the food source again. If however, the animal depletes the food source, the increased frequency of high-angled turns would cause the animal to search the local environment efficiently for new food. Upon failing to encounter new food in the local environment, the frequency of the high-angled turns diminishes and the animal then explores for more distant sources of food.
Dopamine signaling also allows C. elegans to alter their behavior based upon previous experience. That is, dopamine affects learning in C. elegans. For example, wild-type animals respond to a nonlocalized mechanical stimulus (plate tapping) by either backing or by accelerating their forward locomotion rate (Rose and Rankin, 2001). Upon repeated tapping of the plate wild-type animals habituate to the stimulus and display a decreased frequency of reversals. Habituation to plate tapping is modulated by dopamine signaling as cat-2 mutants and mutants of the D1-like dopamine receptor dop-1 habituate to tap more rapidly than do wild-type animals. Additionally, normal tap response can be restored in cat-2 mutants by exposing them to exogenous dopamine (Sanyal et al., 2004). Interestingly dopamine does not control whether or not the animal can respond or habituate to tap as both wild-type and mutant animals respond to repeated tapping by reducing reversal rates to the same extent, rather dopamine affects how rapidly the animal responds to repeated tapping. DOP-1 is expressed in the mechanosensory neurons ALM and PLM (among others) and rescue experiments showed that dopamine acts through the DOP-1 receptor in the mechanosensory neurons to modulate the tap response (Sanyal et al., 2004). The molecular mechanisms used by dopamine to modulate habituation remain unclear but could certainly be limited to the modulation of the activity or the recovery from activation of the ALM and PLM neurons.
Dopamine is also required for another form of learning, state-dependent olfactory adaptation (see Chemosensation in C. elegans). Animals exposed for prolonged periods to an odorant adapt to the odorant stimulus and exhibit a diminished response to the odorant upon reexposure. Such adaptation lasts for several hours (Colbert and Bargmann, 1995). Wild-type animals adapted to an odorant while intoxicated by ethanol exhibit adaptation to the odorant only if ethanol is again present during reexposure. If such adapted animals are instead reexposed to the odorant in the absence of intoxicating concentrations of ethanol they respond to the odorant as if they had not been preexposed. Thus adaptation to the odorant is state-dependent. The ability to acquire state-dependent adaptation requires dopamine as cat-1 and cat-2 mutants adapted to an odorant in the presence of ethanol show adaptation regardless of whether ethanol is present during the testing period (Bettinger and McIntire, 2004). The signaling mechanisms through which dopamine effects state-dependent learning have not yet been identified.
Four dopamine receptors (DOP-1 through DOP-4) have been identified in C. elegans, including homologs of each of the two classes of dopamine receptor (D1- and D2-like) found in mammals (see Table 2). Mutants are available in each receptor and analysis of the receptor mutants has allowed assignment of particular receptors to the modulation of specific behaviors. For example, mutants in the D2-like dopamine receptor DOP-3 fail to slow in response to a bacterial lawn. Thus, DOP-3 is the receptor activated by dopamine in response to a bacterial lawn to slow locomotion rate. The slowing defect of dop-3 mutants is rescued by mutations in the D1-like receptor DOP-1 indicating that DOP-1 and DOP-3 antagonize each other to control the rate of locomotion in response to changing environmental stimuli (Chase et al., 2004). In C. elegans then, as in mammals, D1- and D2-like receptors can function antagonistically to control behavior. The DOP-1 and DOP-3 receptors are coexpressed in the motor neurons of the ventral cord and function there to modulate locomotion rate. The ventral cord motor neurons are not postsynaptic to the dopaminergic neurons, and thus dopamine can function extrasynaptically in C. elegans. As mentioned earlier, mutations in the DOP-1 receptor also cause defects in the ability of animals to respond to mechanical stimulation (plate tapping). DOP-1 functions in the touch neurons to modulate this response, and these neurons are also not postsynaptic to the dopaminergic neurons.
Dopamine may control other behaviors in addition to those revealed by the analysis of cat-2 mutants, as cat-2 null mutants retain significant levels of dopamine (Sanyal et al., 2004). In mammals, enzymes known as tyrosinases may be capable of converting tyrosine to L-DOPA to partially bypass the requirement for tyrosine hydroxylase (Rios et al., 1999). Four putative tyrosinases (tyr-1 through tyr-4) have been identified in C. elegans. Elimination of all dopamine signaling may be possible by combining mutations in the tyrosinases with cat-2 mutations and the analysis of such mutants may reveal additional behaviors controlled by dopamine signaling not observed previously in the analysis of cat-2 mutants.
Serotonin is made in eight types of neurons in C. elegans (see Table 1). Its role in modulating behavior has been established largely through analysis of mutants for the genes encoding the serotonin biosynthetic enzymes, or through behavioral studies of wild-type animals exposed to drugs thought to alter serotonin signaling. Treatment with exogenous serotonin causes dramatic behavioral effects: it inhibits locomotion and the defecation motor program, but stimulates egg laying and pharyngeal pumping (Weinshenker et al., 1995; Horvitz et al., 1982; Segalat et al., 1995; Sawin et al., 2000; Rogers et al., 2001; Niacaris and Avery, 2003; Waggoner et al., 1998; Mendel et al., 1995). Defects in some of the same behaviors are observed in mutants that lack activity of the serotonin biosynthetic enzymes BAS-1 or CAT-4 (Loer and Kenyon, 1993; Waggoner et al., 1998; Sawin et al., 2000; Figure 1).
Serotonin signaling allows C. elegans to respond to changes in its environment by modulating locomotion behavior. Food-deprived animals slow their locomotion rate dramatically when they encounter a bacterial lawn and this “enhanced slowing response” requires serotonin as bas-1 and cat-4 mutants (but not cat-2 mutants) exhibit defects in enhanced slowing (Sawin et al., 2000). Furthermore, treatment of bas-1; cat-4 double mutants with exogenous serotonin can rescue the enhanced slowing defects. The enhanced slowing response caused by serotonin signaling is much more dramatic than the slowing response caused by dopamine signaling, as many food-deprived animals come to a full stop as they encounter food whereas well-fed animals often simply slow their locomotion rate. Serotonin signaling thus provides a mechanism to ensure that food-deprived animals absolutely do not leave a food source once they have encountered it whereas dopamine signaling in well-fed animals encourages the animal to stay in the proximity of food but still may permit limited exploration for new or better food sources.
Serotonin is synthesized in the neurosecretory motor neurons (NSMs), which have sensory endings in the lumen of the pharynx that might sense food, and which also have access to the outside of the pharynx and the pseudocoelom, where serotonin might be released to communicate with the rest of the animal. Thus serotonergic neurons are well positioned to signal a favorable food environment and allow the animal to remain in the area of food to feed and to have progeny. Recently, a mutation was isolated in the tph-1 gene, which encodes another serotonin biosynthetic enzyme, tryptophan hydroxylase (see Figure 1). tph-1 mutants show a modest reduction in egg laying and a reduction in the frequency of pharyngeal pumping, but have not yet been tested for defects in defecation or locomotion (Sze et al., 2000).
Four receptors have been identified that bind serotonin, including three G protein coupled (metabotropic) receptors (SER-1, SER-4 and SER-7) and one serotonin-gated chloride channel (MOD-1; Table 2; Olde and McCombie, 1997; Hamdan et al., 1999; Hobson et al., 2003; Ranganathan et al., 2000). Analysis of mutants for these receptors has shed light on the mechanisms by which serotonin modulates locomotion, pharyngeal pumping, and egg laying.
The serotonin-gated chloride channel, MOD-1, appears to be required for enhanced slowing, but cells expressing MOD-1 have not been determined (Sawin et al., 2000; Ranganathan et al., 2000). Of the three G protein-coupled receptors, SER-1 has the clearest role in the control of egg laying, and might also be responsible for the effects of serotonin on pharyngeal pumping. SER-1 is expressed on the vulval muscles and ser-1 mutants fail to lay eggs in response to serotonin (Dempsey et al., 2005; Carnell et al., 2006). SER-1 is additionally expressed on pharyngeal muscles and may be responsible for the observed effect of serotonin on the repolarization of the pharyngeal muscles (Dempsey et al., 2005; Carnell et al., 2006; Niacaris et al., 2003). SER-4 and SER-7 are also expressed in pharyngeal neurons or muscles and so might also play a role in controlling pharyngeal activity (Hobson et al., 2003; Tsalik et al., 2003). While MOD-1, SER-1, SER-4 and SER-7 are the only serotonin receptors for which pharmacological criteria are available, the expression patterns of several other putative serotonin receptors have been examined (Carre-Pierrat et al., 2006).
Serotonin signaling has been studied extensively in the control of egg laying (see Egg laying). Serotonin is made in two classes of neurons that directly innervate the vulval muscles: the VC4/5 neurons (Duerr et al., 1999) and the pair of HSN hermaphrodite-specific neurons (Desai et al., 1988; McIntire et al., 1992; Sze et al., 2000). A strong case has been made that HSN-released serotonin stimulates egg laying. Exogenous serotonin causes egg laying, and mutants in which the HSN neurons die (egl-1 mutants) are strongly egg-laying defective (Egl). Selective serotonin-reuptake inhibitors, which cause endogenous serotonin to persist in synapses after release, cause egg laying in wild-type animals but not in egl-1 mutants, suggesting the reuptake inhibitors potentiate the effects of HSN-released serotonin to stimulate egg laying. However, serotonin release is not solely responsible for the stimulation of egg laying, as mutations in the serotonin biosynthetic enzymes BAS-1 and CAT-4 do not cause animals to become Egl, and tph-1 mutants are only modestly defective in egg laying (Weinshenker et al., 1995; Sze et al., 2000). Thus, other neurotransmitters released from the HSN neurons must function together with serotonin to stimulate egg laying. In addition to stimulating egg laying, serotonin is also thought to have an inhibitory effect on egg laying: calcium imaging studies suggest that serotonin autoreceptors inhibit HSN activity (Shyn et al., 2003). One model is that serotonin release from the HSN stimulates a cluster of egg laying events, but also inhibits further HSN activity, explaining the ~20 minute interval between clusters of egg-laying events that typically occurs (Waggoner et al., 1998).
Table 3. Drugs and their effects on C. elegans
| Drug | Mammalian / invertebrate targets | C. elegans targetsa | Effects of drugs on wild-type C. elegans |
|---|---|---|---|
| Dopamine | Dopamine receptors | DOP-1, DOP-310, 28 | Inhibits locomotion1, 13, 10. Inhibits egg laying1, 2. Inhibits defecation2, 13. Increases frequency of high-angled turns25. |
| Serotonin | Serotonin receptors | SER-126, 27, SER-226, 5, MOD-517 MOD-113, 14 | Inhibits locomotion11, 3, 13. Inhibits defecation2, 11. Stimulates pharyngeal pumping11, 3, 8, 6 (in the absence of food). Stimulates egg laying2, 3, 7, 12. Causes male tail curling4. Causes reduced sensitivity to aldicarb-induced paralysis21. Stimulates vulval muscle calcium transients22. Inhibits HSN calcium transients22. Inhibits olfactory adaptation24. |
| Octopamine | Octopamine receptors | Causes uncoordinated locomotion3. Inhibits pharyngeal pumping3, 8, 6 (in the presence of food or 5-HT). Inhibits egg laying3, 11 (in the presence of food or 5-HT). Inhibits defecation2, 3. | |
| Tyramine | Tyramine receptors | SER-25 | Inhibits egg laying (5, 9) (in the presence of food or 5-HT). |
| Quinpirole | Dopamine receptors (agonist) | Inhibits locomotion11. Inhibits egg laying11. | |
| Chlorpromazine | D2 receptors (antagonist) | Stimulates egg laying2. | |
| Raclopride | Dopamine receptors (antagonist) | Blocks the effects of dopamine on turning frequency25. Eliminates area-restricted search behavior25 | |
| α-methyl 5-HT | Serotonin receptors (agonist) | SER-129 | Stimulates egg laying23. |
| Methiothepin | Serotonin receptors (antagonist) | MOD-114 | Blocks 5-HT-induced inhibition of locomotion13. Causes defects in enhanced slowing13. Causes hypersensitivity to aldicarb-induced paralysis21. |
| Mianserin | Serotonin receptors (antagonist) | MOD-114 | Blocks 5-HT-induced inhibition of locomotion13. Causes defects in enhanced slowing13. |
| Gramine | Serotonin receptors (antagonist) | Inhibits pharyngeal pumping8. | |
| Ketanserin | Serotonin receptors (antagonist) | Causes hypersensitivity to aldicarb-induced paralysis21. | |
| Fluoxetine | Serotonin-specific reuptake transporter (inhibitor) | MOD-5 and others16, 17, 26 | Stimulates egg laying2, 16. Potentiates enhanced slowing response13, 17. Stimulates nose contraction16. Inhibits locomotion16, 21. |
| Paroxetine | Serotonin-specific reuptake transporter (inhibitor) | Stimulates nose contraction16. | |
| Clomipramine | Serotonin reuptake transporter (inhibitor) | Stimulates egg laying2. Stimulates nose contraction16. | |
| Imipramine | Monoamine reuptake transporter (inhibitor) | EGL-218 | Stimulates egg laying2, 12, 19. Stimulates pharyngeal pumping20. |
| Reserpine | Monoamine vesicular transporter (inhibitor) | Stimulates locomotion15 (in the presence of food). Inhibits egg laying15. | |
| Phentolamine | Octopamine and adrenergic receptors (antagonist) | Stimulates egg laying3. | |
| aTo qualify as a C. elegans target for a particular drug either: 1) mutation of the gene encoding the target protein must cause amine or drug-specific behavioral defects and lessen the behavioral effects of exogenous drug exposure; or 2) the target protein must bind to the drug in vitro and mutation of the gene encoding the target protein must lessen the effects of the exogenous drug in vivo. | |||
References
| 1. Schafer and Kenyon, 1995 |
| 2. Weinshenker et al., 1995 |
| 3. Horvitz et al., 1982 |
| 4. Loer and Kenyon, 1993 |
| 5. Rex et al., 2004 |
| 6. Rogers et al., 2001 |
| 7. Waggoner et al., 1998 |
| 8. Niacaris and Avery, 2003 |
| 9. Alkema et al., 2005 |
| 10. Chase et al., 2004 |
| 11. Segalat et al., 1995 |
| 12. Mendel et al., 1995 |
| 13. Sawin et al., 2000 |
| 14. Ranganathan et al., 2000 |
| 15. Duerr et al., 1999 |
| 16. Choy and Thomas, 1999 |
| 17. Ranganathan et al., 2001 |
| 18. Weinshenker et al., 1999 |
| 19. Trent et al., 1983 |
| 20. Avery and Horvitz, 1990 |
| 21. Nurrish et al., 1999 |
| 22. Shyn et al., 2003 |
| 23. Bastiani et al., 2003 |
| 24. Nuttley et al., 2002 |
| 25. Hills et al., 2004 |
| 26. Dempsey et al., 2005 |
| 27. Hamdan et al., 1999 |
| 28. Suo et al., 2002 |
| 29. Carnell et al., 2006 |
DLC was funded by the National Institutes of Health grant MH077935-01, the National Parkinson Foundation and the Parkinson Disease Foundation. MRK was funded by National Institutes of Health grant NS036918.
Alkema, M.J., Hunter-Ensor, M., Ringstad, N., Horvitz, H.R. (2005). Tyramine Functions Independently of Octopamine in the Caenorhabditis elegans Nervous System. Neuron 46, 247–260. Abstract Article
Avery, L., and Horvitz, H.R. (1990). Effects of starvation and neuroactive drugs on feeding in Caenorhabditis elegans. J. Exp. Zool. 253, 263–270. Abstract Article
Bargmann, C.I. (1998). Neurobiology of the Caenorhabditis elegans genome. Science 282, 2028–2033. Abstract Article
Bastiani, C.A., Gharib, S., Simon, M.I., Sternberg, P.W. (2003). Caenorhabditis elegans Galphaq regulates egg-laying behavior via a PLCbeta-independent and serotonin-dependent signaling pathway and likely functions both in the nervous system and in muscle. Genetics 165, 1805–1822. Abstract Article
Bettinger, J.C., and McIntire, S.L. (2004). State-dependency in C. elegans. Genes Brain Behav. 3, 266–272. Abstract Article
Carnell L, Illi J, Hong SW, McIntire SL. (2006). The G-protein-coupled serotonin receptor SER-1 regulates egg laying and male mating behaviors in Caenorhabditis elegans. J. Neurosci. 25, 10671–10681. Abstract Article
Carre-Pierrat M, Baillie D, Johnsen R, Hyde R, Hart A, Granger L, Segalat L. (2006). Characterization of the Caenorhabditis elegans G protein-coupled serotonin receptors. Invert. Neurosci. 6, 189–205. Abstract Article
Chase, D.L., Pepper, J.S., Koelle, M.R. (2004). Mechanism of extrasynaptic dopamine signaling in Caenorhabditis elegans. Nat. Neurosci. 7, 1096–1103. Abstract Article
Choy, R.K., Thomas, J.H. (1999). Fluoxetine-resistant mutants in C. elegans define a novel family of transmembrane proteins. Mol. Cell 4, 143–152. Abstract Article
Colbert, H.A., Bargmann, C.I. (1995). Odorant-specific adaptation pathways generate olfactory plasticity in C. elegans. Neuron 14, 803–812. Abstract Article
Dempsey, C.M., Mackenzie, S.M., Gargus, A., Blanco, G., Sze, J.Y. (2005). Serotonin (5HT), Fluoxetine, Imipramine and Dopamine Target Distinct 5HT Receptor Signaling to Modulate Caenorhabditis elegans Egg-Laying Behavior. Genetics 169, 1425–1436. Abstract Article
Desai, C., Garriga, G., McIntire, S.L., Horvitz, H.R. (1988). A genetic pathway for the development of the Caenorhabditis elegans HSN motor neurons. Nature 336, 638–646. Abstract Article
Duerr, J.S., Frisby, D.L., Gaskin, J., Duke, A., Asermely, K., Huddleston, D., Eiden, L.E., Rand, J.B. (1999). The cat-1 gene of Caenorhabditis elegans encodes a vesicular monoamine transporter required for specific monoamine-dependent behaviors. J. Neurosci. 19, 72–84. Abstract Article
Hamdan, F.F., Ungrin, M.D., Abramovitz, M., Ribeiro, P. (1999). Characterization of a novel serotonin receptor from Caenorhabditis elegans: cloning and expression of two splice variants. J. Neurochem. 72, 1372–1383. Abstract Article
Hills, T., Brockie, P.J., Maricq, A.V. (2004). Dopamine and glutamate control area-restricted search behavior in Caenorhabditis elegans. J. Neurosci. 24, 1217–1225. Abstract Article
Hobson, R.J., Geng, J., Gray, A.D., Komuniecki, R.W. (2003). SER-7b, a constitutively active Galphas coupled 5-HT7-like receptor expressed in the Caenorhabditis elegans M4 pharyngeal motorneuron. J. Neurochem. 87, 22–29. Abstract Article
Hobson, R.J., Hapiak, V.M., Xiao, H., Buehrer, K.L., Komuniecki, P.R., Komuniecki, R.W. (2006). SER-7, a Caenorhabditis elegans 5-HT7-like receptor, is essential for the 5-HT stimulation of pharyngeal pumping and egg laying. Genetics 172, 159–169. Abstract Article
Horvitz, H.R., Chalfie, M., Trent, C., Sulston, J.E., Evans, P.D. (1982). Serotonin and octopamine in the nematode Caenorhabditis elegans. Science 216, 1012–1014. Abstract Article
Komuniecki, R.W., Hobson, R.J., Rex, E.B., Hapiak, V.M., Komuniecki, P.R. (2004). Biogenic amine receptors in parasitic nematodes: what can be learned from Caenorhabditis elegans? Mol. Biochem. Parasitol. 137, 1–11. Abstract Article
Lints, R., and Emmons, S.W. (1999). Patterning of dopaminergic neurotransmitter identity among Caenorhabditis elegans ray sensory neurons by a TGFbeta family signaling pathway and a Hox gene. Development 126, 5819–5831. Abstract Article
Liu, K.S., and Sternberg, P.W. (1995). Sensory regulation of male mating behavior in Caenorhabditis elegans. Neuron 14, 79–89. Abstract Article
Loer, C.M., and Kenyon, C.J. (1993). Serotonin-deficient mutants and male mating behavior in the nematode Caenorhabditis elegans. J. Neurosci. 13, 5407–5417. Abstract Article
Lynch, A.S., Briggs, D., Hope, I.A. (1995). Developmental expression pattern screen for genes predicted in the C. elegans genome sequencing project. Nat. Genet. 11, 309–313. Abstract Article
McIntire, S.L., Garriga, G., White, J., Jacobson, D., Horvitz, H.R. (1992). Genes necessary for directed axonal elongation or fasciculation in C. elegans. Neuron 8, 307–322. Abstract Article
Mendel, J.E., Korswagen, H.C., Liu, K.S., Hajdu-Cronin, Y.M., Simon, M.I., Plasterk, R.H., Sternberg, P.W. (1995). Participation of the protein Go in multiple aspects of behavior in C. elegans. Science 267, 1652–1655. Abstract Article
Nass, R., Hall, D.H., Miller, D.M. 3rd, Blakely, R.D. (2002). Neurotoxin-induced degeneration of dopamine neurons in Caenorhabditis elegans. Proc. Natl. Acad. Sci. U.S.A. 99, 3264–3269. Abstract Article
Niacaris, T., and Avery, L. (2003). Serotonin regulates repolarization of the C. elegans pharyngeal muscle. J. Exp. Biol. 206, 223–231. Abstract Article
Nurrish, S., Segalat, L., Kaplan, J.M. (1999). Serotonin inhibition of synaptic transmission: Galpha(0) decreases the abundance of UNC-13 at release sites. Neuron 24, 231–242. Abstract Article
Nuttley, W.M., Atkinson-Leadbeater, K.P., Van Der Kooy, D. (2002). Serotonin mediates food-odor associative learning in the nematode Caenorhabditis elegans. Proc. Natl. Acad. Sci. U.S.A. 99, 12449–12454. Abstract Article
Olde, B., and McCombie, W.R. (1997). Molecular cloning and functional expression of a serotonin receptor from Caenorhabditis elegans. J. Mol. Neurosci. 8, 53–62. Abstract
Ranganathan, R., Cannon, S.C., Horvitz, H.R. (2000). MOD-1 is a serotonin-gated chloride channel that modulates locomotory behaviour in C. elegans. Nature 408, 470–475. Abstract Article
Ranganathan, R., Sawin, E.R., Trent, C., Horvitz, H.R. (2001). Mutations in the Caenorhabditis elegans serotonin reuptake transporter MOD-5 reveal serotonin-dependent and -independent activities of fluoxetine. J. Neurosci. 21, 5871–5884. Abstract Article
Rex, E., Hapiak, V., Hobson, R., Smith, K., Xiao, H., and Komuniecki, R. (2005). TYRA-2 (F01E11.5): a Caenorhabditis elegans tyramine receptor expressed in the MC and NSM pharyngeal neurons. J. Neurochem. 94, 181–191. Abstract Article
Rex, E., Molitor, S.C., Hapiak, V., Xiao, H., Henderson, M., Komuniecki, R. (2004). Tyramine receptor (SER-2) isoforms are involved in the regulation of pharyngeal pumping and foraging behavior in Caenorhabditis elegans. J. Neurochem. 91, 1104–1115. Abstract Article
Rex, E., Komuniecki, R.W. (2002). Characterization of a tyramine receptor from Caenorhabditis elegans. J. Neurochem. 82, 1352–1359. Abstract Article
Rios, M., Habecker, B., Sasaoka, T., Eisenhofer, G., Tian, H., Landis, S., Chikaraishi, D., and Roffler-Tarlov, S. (1999). Catecholamine synthesis is mediated by tyrosinase in the absence of tyrosine hydroxylase. J. Neurosci. 19, 3519–3526. Abstract Article
Rogers, C.M., Franks, C.J., Walker, R.J., Burke, J.F., Holden-Dye, L. (2001). Regulation of the pharynx of Caenorhabditis elegans by 5-HT, octopamine, and FMRFamide-like neuropeptides. J. Neurobiol. 49, 235–244. Abstract Article
Rose, J.K., and Rankin, C.H. (2001). Analyses of habituation in Caenorhabditis elegans. Learn Mem. 8, 63–99. Abstract Article
Sanyal, S., Wintle, R.F., Kindt, K.S., Nuttley, W.M., Arvan, R., Fitzmaurice, P., Bigras, E., Merz, D.C., Hebert, T.E., van der Kooy, D., Schafer, W.R., Culotti, J.G., Van Tol, H.H. (2004). Dopamine modulates the plasticity of mechanosensory responses in Caenorhabditis elegans. EMBO J. 23, 473–482. Abstract Article
Sawin, E.R., Ranganathan, R., Horvitz, H.R. (2000). C. elegans locomotory rate is modulated by the environment through a dopaminergic pathway and by experience through a serotonergic pathway. Neuron 26, 619–631. Abstract Article
Schafer, W.R., and Kenyon, C.J. (1995). A calcium-channel homologue required for adaptation to dopamine and serotonin in Caenorhabditis elegans. Nature. 375,73–78. Abstract Article
Segalat, L., Elkes, D.A., Kaplan, J.M. (1995). Modulation of serotonin-controlled behaviors by Go in Caenorhabditis elegans. Science 267, 1648–1651. Abstract Article
Shyn, S.I., Kerr, R., Schafer, W.R. (2003). Serotonin and Go modulate functional states of neurons and muscles controlling C. elegans egg-laying behavior. Curr. Biol. 13, 1910–1915. Abstract Article
Sugiura, M., Fuke, S., Suo, S., Sasagawa, N., Van Tol, H.H., and Ishiura, S. (2005). Characterization of a novel D2-like dopamine receptor with a truncated splice variant and a D1-like dopamine receptor unique to invertebrates from Caenorhabditis elegans. J. Neurochem. 94, 1146–1157. Abstract Article
Sulston, J., Dew, M., Brenner, S. (1975). Dopaminergic neurons in the nematode Caenorhabditis elegans. J. Comp. Neur. 163, 215–226. Abstract Article
Sulston, J.E., and Horvitz, HR. (1977). Post-embryonic cell lineages of the nematode, Caenorhabditis elegans. Dev. Biol. 56, 110–156. Abstract Article
Suo, S., Sasagawa, N., Ishiura, S. (2002). Identification of a dopamine receptor from Caenorhabditis elegans. Neurosci. Lett. 319, 13–16. Abstract Article
Suo, S., Sasagawa, N., Ishiura, S. (2003). Cloning and characterization of a Caenorhabditis elegans D2-like dopamine receptor. J. Neurochem. 86, 869–878. Abstract Article
Suo, S., Kimura, Y., Van Tol, H.H.M. (2006). Starvation induces cAMP response element-binding protein-dependent gene expression through octopamine-Gq signaling in Caenorhabditis elegans. J. Neurosci. 26, 10082–10090. Abstract Article
Sze, J.Y., Victor, M., Loer, C., Shi, Y., Ruvkun, G. (2000). Food and metabolic signalling defects in a Caenorhabditis elegans serotonin-synthesis mutant. Nature 403, 560–564. Abstract Article
Trent, C., Tsuing, N., Horvitz, H.R. (1983). Egg-laying defective mutants of the nematode Caenorhabditis elegans. Genetics 104, 619–647. Abstract Article
Tsalik, E.L., Niacaris, T., Wenick, A.S., Pau, K., Avery, L., Hobert, O. (2003). LIM homeobox gene-dependent expression of biogenic amine receptors in restricted regions of the C. elegans nervous system. Dev. Biol. 263, 81–102. Abstract Article
Waggoner, L.E., Zhou, G.T., Schafer, R.W., Schafer, W.R. (1998). Control of alternative behavioral states by serotonin in Caenorhabditis elegans. Neuron 21, 203–214. Abstract Article
*Edited by Erik M. Jorgensen and Joshua M. Kaplan. Last revised December 15, 2006. Published February 20, 2007. This chapter should be cited as: Chase, D.L. and Koelle, M.R. Biogenic amine neurotransmitters in C. elegans (February 20, 2007), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.132.1, http://www.wormbook.org.
Copyright: © 2007 Daniel L. Chase and Michael R. Koelle. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
§To whom correspondence should be addressed. E-mail: danchase@biochem.umass.edu
 All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.
All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.