Embryo series courtesy of Einhard Schierenberg
Embryo series courtesy of Einhard SchierenbergTable of Contents
Abstract
Synaptogenesis is a process involving the formation of a neurotransmitter release site in the presynaptic neuron and a receptive field at the postsynaptic partners, and the precise alignment of pre- and post-synaptic specializations. In C. elegans synapses are found as en passant axonal swellings along the nerve processes. Genetic screens using a synaptic vesicle-associated GFP marker have identified key players in synaptic target recognition and organization of the presynaptic terminals. Importantly, the functions of most genes are evolutionarily conserved. Further studies using a combination of genetic modifier screens and reverse genetics have begun to reveal the underlying signaling pathways.
Synapses are morphologically distinct subcellular junctional structures, composed of a presynaptic terminal, a postsynaptic target and the synaptic cleft aligning pre- and post-synaptic specializations (Cowan et al., 2001; Pappas and Purpura, 1972). The presynaptic terminal is characterized by a cluster of synaptic vesicles surrounding the electron-dense membrane specializations; and the postsynaptic site contains densely packed ion channels and signal transduction molecules. Both the pre- and post-synaptic specializations display variable appearances depending on the organisms and neuronal types (De Camilli et al., 2001; Sorra and Harris, 2000; Zhai and Bellen, 2004).
Synapse formation occurs in two general manners: en passant—synaptic boutons are formed along the axon shaft; or terminaux—synaptic boutons are formed at the end of axon branches. C. elegans synapses occur en passant between neighbouring parallel nerve processes, or between nerve processes and muscle arms (White et al., 1986). The presynaptic site is seen as a vesicle-filled varicosity containing the electron-dense presynaptic density in the membrane adjacent to the contact with the postsynaptic elements (Figure 1). On samples prepared by traditional fixation method clear and spherical synaptic vesicles (SV) are usually 35–45 nm in diameter, and large dense-core vesicles (LDCV) range from 40–53nm. The size of the presynaptic regions, defined by the axonal swellings and the number of SV and the size of density, varies considerably, even among the same types of synapses or among the synapses from the same neuron. The prominence of presynaptic density also varies in a way that does not necessarily correspond to the number of vesicles. The overall ultrastructural appearance of presynaptic terminals of worm synapses bears much resemblance to those of many synapses in the vertebrate central nervous system. However, the postsynaptic sites of worm synapses usually have no visible density-like structures that are commonly found in other organisms. Therefore, synaptic partners are assigned mostly based on proximity, with monad synapse having one postsynaptic partner, dyad with two, and triad with three postsynaptic partners. In the case of dyad and triad synapses, it is difficult to tell whether all partners are functional. The presence of more than one postsynaptic target also raises the possibility that synaptogenesis may depend on the simultaneous presence of two particular postsynaptic elements. The number of synapses each nerve process makes is quite variable (White et al., 1986). Designated synaptic partners often make contact through fasciculation for a long distance, but form synapses only at specific sites. This chapter focuses on presynaptic assembly and synaptic target selection. For more information on postsynaptic assembly, see Synaptic function.
A key reagent to examine synapses in vivo is the synaptobrevin GFP fusion reporter (Nonet, 1999). Synaptobrevin (SNB-1/T0H9.4) is a 109 amino acid, integral membrane protein of synaptic vesicles. The original SNB-1::GFP marker has GFP fused at the C-terminal of SNB-1, and by topology prediction, GFP would reside inside the vesicle. Evidence for this marker as labeling synaptic vesicles are (1) colocalization of SNB-1::GFP with endogenous SNB-1 by wholemount immunocytochemistry; (2) SNB-1::GFP fluorescence is seen predominantly in cell bodies in the unc-104 kinesin mutant; and (3) deletion of the transmembrane domain of SNB-1(Δ TM)::GFP causes GFP to be not restricted to synaptic regions. SNB-1::GFP expressed in interneurons and HSN neurons show close apposition to postsynaptic markers (GLR-1 and LIN-10; Rongo et al., 1998; Shen and Bargmann, 2003). However it is not yet precisely known which population of synaptic vesicles the SNB-1::GFP fluorescence observed represents. A variety of synaptic GFP reporter genes have also recently been developed (Sieburth et al., 2005; Yeh et al., 2005; Francis et al., 2005).
The success of screening mutants for synaptogenesis defects has depended critically on several factors. First, the worm is transparent and GFP is readily observable in living animals. Second, much of the nervous system function is not required for viability or fertility, making it easy to maintain homozygous mutants. Third, expression of SNB-1::GFP under neuron-specific promoters allows the optical resolution of single synapses; and the choice of neurons or synapses seems to be an important factor to the class of mutants isolated. Lastly, visual inspection of SNB-1::GFP pattern following the traditional mutagenesis by far is most rewarding, although it is labor intensive.
Several screens have been performed using SNB-1::GFP to label synaptic vesicle clusters in ASI chemosensory neurons (Pstr-3; Crump et al., 2001), mechanosensory neurons (Pmec-7; Schaefer et al., 2000), type D motor neurons (Punc-25; Zhen and Jin, 1999) and HSN neurons (Punc-86; Shen and Bargmann, 2003; Figure 2), which identified sets of overlapping genes, named sad, sam, syd, and syg, respectively. Most mutations appear to have wide effects on many or all synapses, but cause little behavior abnormalities. Moreover, the terminal synaptic phenotypes differ in a synapse-type specific manner. Importantly, although the mutations were isolated based on abnormal patterns of SNB-1::GFP, the molecular characterizations thus far have shown that none of the genes is a component of synaptic vesicles, rather these genes define distinct subdomains within presynaptic terminals.
|
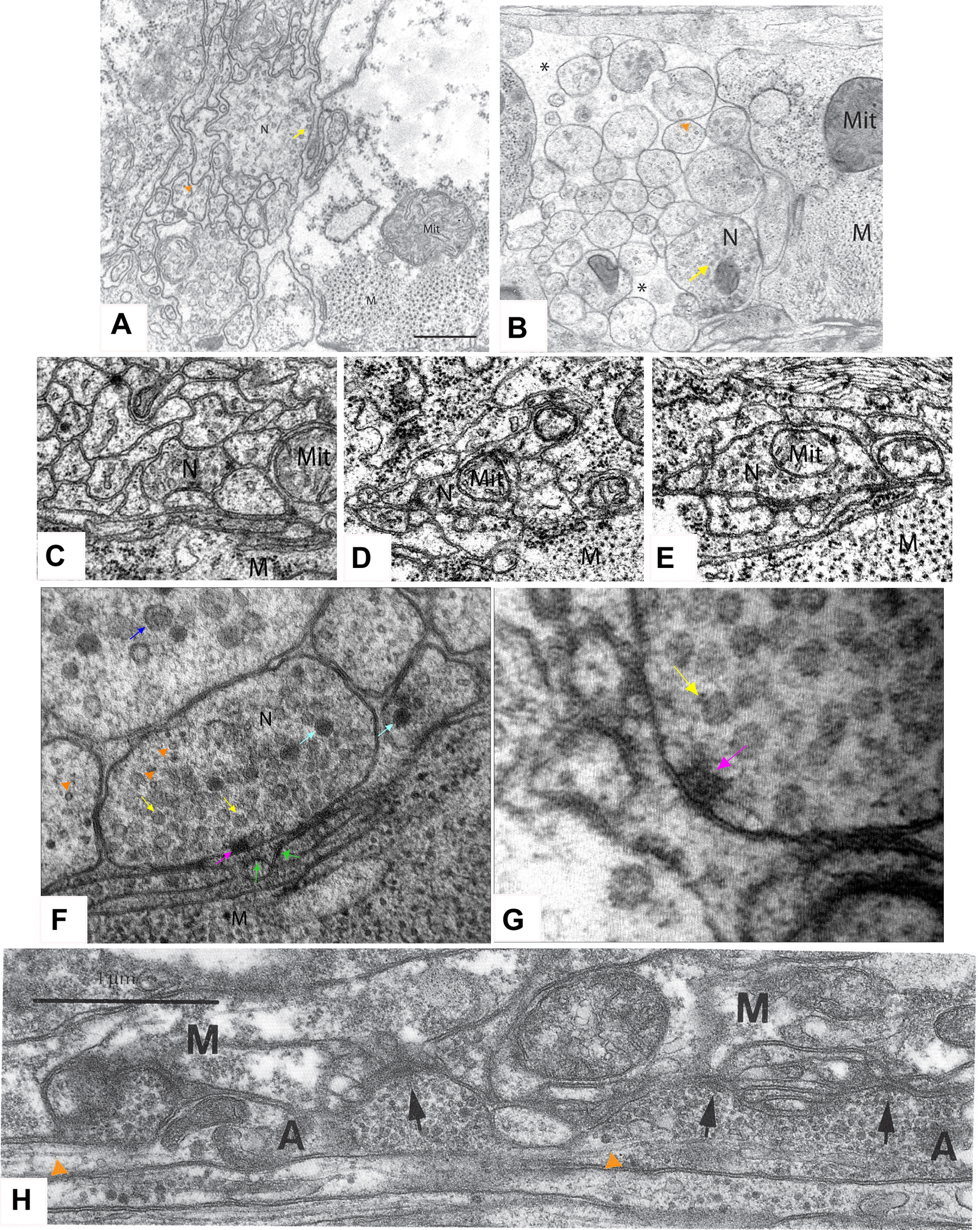
Figure 1. Electron microscope images of worm synapses. (A) Ventral nerve cord fixed by classical method using osmium and glutaraldehyde. (B) Ventral nerve cord fixed by high-pressure freezing. Images in A and B are taken from (Rostaing et al., 2004) with permission. M, muscle; N, presynaptic terminal; Mit, mitochondria. Yellow arrows, SV; orange triangles, MT; *, space between nerve processes. Scale: 500nm. (C–E) Images of synapses by classical fixation method: neuron-neuron synapse in the ventral nerve cord (C), cholinergic neuromuscular junction (D), and GABAergic NMJ (E). M, muscle; N, presynaptic terminal; Mit, mitochondria. (F–G). Images of synapse by high-pressure fixation procedure. G shows an enlarged view of the presynaptic density (R. Weimer and J-L. Bessereau, personal communication). Yellow arrows, SV; light-blue arrows, dense core vesicles; dark-blue arrow, large core vesicle; green arrows, postsynaptic density-like structures; pink-purple arrow, presynaptic density; orange triangle, MT. (H). Image of longitudinal section of ventral nerve cord, showing the sharp transition of SV cluster and MT nearby. Black arrows, presynaptic density; orange triangles, MT; A, axon; M, muscle. Taken from Hedgecock and Hall (1991) with permission.
Presynaptic assembly involves coordinated action of several inter-dependent events (Murthy and De Camilli, 2003): formation of the electron-dense presynaptic density at the membrane apposition to the post-synaptic cell; clustering of the synaptic vesicles surrounding the density; and establishment of the active zone with the proper ratio of docked vesicles to presynaptic density.
Presynaptic density is in the center of a presynaptic terminal, and is often called “active zone”. Its electron-dense appearance likely reflects the presence of a protein scaffold, frequently referred as “the presynaptic grid” within which vesicles are docked (Akert et al., 1972). The density in worm synapses is simple and small (Figure 1), and does not appear to have elaborate sub-ultrastructural characteristics described for those in large synapses in other animals. It is assumed that the molecular components of presynaptic density include voltage-gated calcium channels–like candidates are UNC-2 and UNC-36 (Tam et al., 2000), SV fusion complex UNC-64/Syntaxin (Saifee et al., 1998) and affiliated proteins that function in vesicle docking and priming such as UNC-10/Rim (Koushika et al., 2001) and UNC-13/mUNC13 (Patton et al., 1997). Several mammalian presynaptic density proteins such as Bassoon and Picoloo (Ziv and Garner, 2004) do not seem to have direct homologs in C. elegans by the worm genome sequence annotation.
|
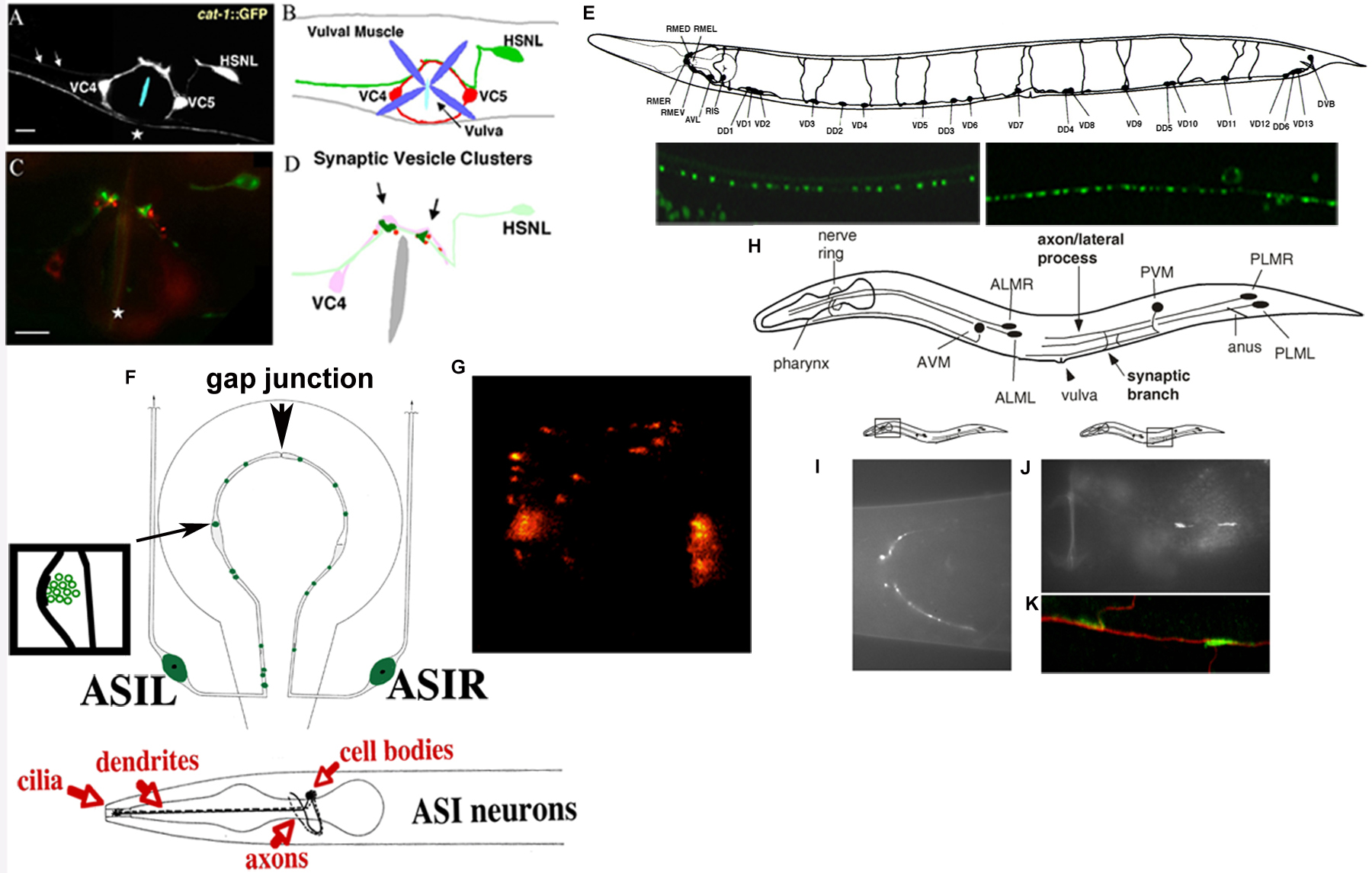
Figure 2. Patterns of SNB-1::GFP in selected neurons. (A–D). HSN synapses visualized by Punc-86SNB-1::YFP. Images are from (Shen and Bargmann, 2003) with permission. (A). Ventral view of an animal expression a cat-1::GFP transgene in HSNL, VC4 and VC5. Arrows point to the segment of HSNL axon anterior to the vulva. (B). Schematic representation of (A). Vulval muscles are included. Anterior is to the left and ventral right is down. (C). Ventral view of a kyIs235 animal with Punc-86SNB-1::YFP colored in green and Punc-4LIN-10::dsRED colored in red. Synaptic vesicles are clutered near the vulval area. (D). Schematic representation of (C). Arrows point to synaptic vesicle clusters. Anterior is to the left and right is down. (E). Synapses of the type D motor neurons, visualized by Punc-25SNB-1::GFP. The D type neurons include six DD and 13 VD neurons. DD form synapses to the dorsal body muscles and VD form synapses to the ventral muscles. Blobs, synapses to muscles; arrows, synaptic inputs to the D neurons. Shown below are images of Punc-25SNB-1::GFP (juIs1) in the dorsal and ventral cords, respectively. (F–G). ASI synapses visualized by Pstr-3SNB-1::GFP. Images are from (Crump et al., 2001) with permission. (F). Flatterned cross-section showing the predicted presynaptic specializations of the ASI neurons (White et al., 1986). The two larger shaded ovals are the cell bodies of ASIL and ASIR. The axons of ASIL and ASIR form gap junctions at their dorsal ends. The smaller shaded circles within the axons represent clusters of synaptic vesicles (enlarged in the insert). (G). Image of Pstr-3SNB-1::GFP (kyIs105) showing the synaptic cluster pattern, the large fluorescent blobs are ASI cell bodies. (H–K). Synapses of the touch neurons, visualized by Pmec-7SNB-1::GFP. Images are from (Schaefer et al., 2000) with permission. (H). The position and morphology of neurons expressing mec-7. (I–J). Pmec-7SNB-1::GFP (jsIs37) images in the head and around vulva. (K). Double images of SNB-1::GFP in green and anti-MEC-7 in red.
Disruptions of vesicle docking, priming and fusion machinery have no obvious effect on the integrity of presynaptic density. Loss of function mutations in unc-64/syntaxin (Saifee et al., 1998), unc-13 (Richmond et al., 1999), and unc-18 (Weimer et al., 2003) each cause a range of defects in synaptic vesicle docking and priming. The UNC-10/Rim (Rab3-interacting-molecule) family of proteins contain conserved Zn-finger, PDZ and C2 domains, and interact with Rab3GTP (Wang et al., 1997), cAMPGEFII (Ozaki et al., 2000), Munc13 (Schoch et al., 2002) and others (Ohtsuka et al., 2002; Wang et al., 2002). In unc-10 mutants, presynaptic density and vesicle docking are normal, and UNC-13 localization is normal (Koushika et al., 2001). There are two cAMPGEFII (pxf-1/T14G10.2 and pxf-2/T20G5.5), but whether they have a role in synapses is not known.
Liprins are conserved proteins containing coiled-coil and SAM domains (Serra-Pages et al., 1998). They were originally identified through binding to the intracellular domain of LAR receptor tyrosine phosphatase, and have now been shown to bind Rim and CAST/ERC (for CAZ-associated structural protein/ELKs-Rab6-interacting protein-CAST; Ko et al., 2003; Schoch et al., 2002). Liprins include two subfamilies, liprin-α and liprin-β, of which only liprins-α bind LAR-RPTP (Serra-Pages et al., 1998). syd-2 encodes the worm liprin-α (Zhen and Jin, 1999). In syd-2 mutants, presynaptic densities are altered and appear less electron-dense than normal. The localization of UNC-10/Rim is less discrete in syd-2 mutants than in wild type (Ackley et al., 2005). Antibody studies show that SYD-2 is localized at synapses (Zhen and Jin, 1999); and immuno-EM puts SYD-2 in the very center of presynaptic density (Yeh et al., 2005). The localization of SYD-2 does not depend on the presence of synaptic vesicles (Zhen and Jin, 1999). elks-1, a worm homolog of CAST/ERC, is localized at synapses; mutations in elks-1 do not cause detectable synaptic defects, and ELKS localization is normal in syd-2 and unc-10 mutants (Deken et al., 2005). These data indicate that SYD-2/Liprin-α is one important organizer of presynaptic density. In vertebrate synapses several isoforms of liprin-α are associated with both presynaptic and postsynaptic densities (Wyszynski et al., 2002). No evidence for a postsynaptic association of SYD-2 has been reported, although SYD-2 is expressed in muscle cells (Zhen and Jin, 1999).
The C. elegans LAR-RPTP, ptp-3, has several isoforms (Harrington et al., 2002). The long isoform PTP-3A most resembles the vertebrate LAR. Antibodies for common region of PTP-3 detects staining in the nervous system; isoform-specific GFP tagged transgenes suggest that PTP-3A is likely the predominant form in synapse (Ackley et al., 2005). The synaptic localization of PTP-3 and SYD-2 shows subtle dependency. Mutations disrupting ptp-3A or ptp-3 show synaptic pattern defects similar to, but weaker than, syd-2(lf). Interestingly, a mutation in nid-1, nidogen, shows similar defects (Ackley et al., 2003). In mammalian cultured cells, LAR can bind Nidogen-laminin (O'Grady et al., 1998). In nid-1 mutants, the synaptic localization of PTP-3 and SYD-2 are both altered. SYD-2 localization is also abnormal in ptp-3 mutants (Ackley et al., 2005). These analyses reveal a functional link between SYD-2, LAR and Nidogen, hinting at the possibility that SYD-2 recruits other density components to facilitate its assembly, and that such presynaptic organizing activity of SYD-2 may be modulated through LAR-Nidogen interaction, which, together with laminin, secure the synaptic assembly.
At synapses, there are multiple forms of vesicles (Bodian, 1972; Murthy and De Camilli, 2003; Figure 1). Biosynthetic precursor vesicles containing synaptic proteins are generated at the cell body and transported to the terminals through MT based motor proteins. The UNC-104 motor was the founding member of the KIF1A kinesins that transport synaptic vesicle precursors (Hall and Hedgecock, 1991). In unc-104 mutant animals, very few mature synaptic vesicles are detected at synapses; whereas small, clear vesicles that resemble synaptic vesicles are retained in the cell bodies. This mutant has been used as a standard genetic background for testing whether proteins are associated with SV. The conventional kinesin-1 is a tetramer composed of heavy chain and light chain, corresponding to UNC-116 and KLC-2 (Patel et al., 1993; Sakamoto et al., 2004; Signor et al., 1999). Mutations in both seem to affect the traffic of SV-like components (Byrd et al., 2001; Sakamoto et al., 2004). EM analysis on unc-116 reveals rather broad and diverse defects in cellular organelle distribution (D. Hall, personal communication), suggesting that kinesin-1 has broad activity in vesicle traffic.
The presence of coated endocytic vesicles at presynaptic membranes indicate that most of the vesicles can be assembled locally from components that either are recycled or are previously transported from the cell soma (Murthy and De Camilli, 2003). C. elegans proteins that function in SV endocytosis such as unc-26/synaptojanin (Harris et al., 2000), unc-57/endophilin (Schuske et al., 2003) and unc-11/AP180 (Nonet et al., 1999; see Synaptic function) are identified based on behavior defects and are mostly identified through aldicarb resistance screens (Nguyen et al., 1995). As expected, these mutants exhibit a severe depletion of SV at synapses. The direct link between the anterograde transported SV precursors and local-recycled SV is not yet known.
In various EM fixation filamentous fibers are seen in synapses (Figure 1; Hedgecock and Hall, 1991; R. Weimer and J-L. Bessereau, personal communication). MT runs very close to, but not within, the synapses. No intermediate filament is identified. Actin is presumed to be concentrated primarily around the active zone and vesicle clusters, and studies in other organisms show that actin likely prevents SV dispersion or to recapture endocytic SV (Goda and Davis, 2003). No direct studies on the role of actins in worm synapses are reported. Spectrins are actin bundling proteins. There are two spectrin genes, unc-70/β-spectrin and spc-1/α-spectrin. The existing mutation in spc-1 causes embryonic lethal (Norman and Moerman, 2002). In unc-70(lf) mutants the number and localization of vesicles are normal, indicating that spectrins are not essential for preserving the vesicle clustering domain (Hammarlund et al., 2000). Synapsins are a family of actin-associated phosphoproteins that are required for maintaining the vesicle reserve pool in vertebrate synapses (Greengard et al., 1994). Worm has a synapsin homolog snn-1/Y38C1BA.2 whose function is not known.
sad-1 mutants were identified through SNB-1::GFP screens for ASI and type D motor neurons (Crump et al., 2001). Ultrastructural analysis shows that synaptic vesicles distribute more broadly than wild type animals, but presynaptic densities are normal. SAD-1 defines a novel conserved family of protein Ser/Thr kinases, with the kinase domain closely related to that of PAR-1 and MARK (MAP/microtubule affinity regulating kinase) family (see Asymmetric cell division and axis formation in the embryo). SAD-1 localization intermingles with SV cluster, but does not depend on synaptic vesicles. Overexpression of SAD-1 can induce clustering of synatic vesicle markers or vesicle precursors at non-synaptic regions. SAD-1 may play a role in organizing synaptic cytoskeleton to pattern vesicle accumulation. In addition, SAD-1 also regulates neuronal polarity as the polarity of the DD neurons is altered in sad-1 mutants (Crump et al., 2001). Studies of mouse SAD-1 homologs reveal a similar role in neuronal polarity (Kishi et al., 2005).
At most synapse, there is an abrupt transition in SV density at the edge of the clusters. The identification of rpm-1 defines a new subsynaptic domain called “Periactive zone”, which is further away from the active zone and devoid of synaptic vesicles and could have a role in separating synaptic domains from non-synaptic cytoplasm. rpm-1 mutants exhibit diverse abnormalities in synaptic morphology. Mechanosensory neurons frequently fail to cluster synaptic vesicles in the proper synaptic sites, and often over-extend axons (Schaefer et al., 2000). Chemosensory neurons have disorganized vesicle clusters (Crump et al., 2001). In motor neurons, EM analysis shows multiple defects including severe reduction of vesicle numbers per synapse, presence of several presynaptic densities within one synaptic terminal and alteration in the ratio of SV to density (Nakata et al., 2005; Zhen et al., 2000).
RPM-1 is a large conserved protein with homologs known as Highwire in Drosophila, Esrom in Zebrafish, Phr1 in mouse and Pam in human. These proteins are all implicated in synapse formation (Burgess et al., 2004; D'Souza et al., 2005; Wan et al., 2000). They have several predicted functional modules, one of which is a Ring finger domain that is a signature of the ubiquitin E3 ligase. An F-box domain containing protein FSN-1 has been identified as RPM-1 binding partner provides, supporting RPM-1 as an E3 ubiquitin-ligase domain (Liao et al., 2004). A genetic suppressor screen of rpm-1 identified three map kinases, the Dual-Leucine-zipper-bearing MAPKKK DLK-1, the MAPKK MKK-4, and the p38-like MAP kinase PMK-3 (Nakata et al., 2005). Inactivation of these three kinases suppresses rpm-1 loss of function phenotypes, whereas elevated expression or constitutive activation of these kinases causes synaptic defects resembling rpm-1(lf). This MAP kinase pathway functions presynaptically. DLK-1 is localized to the periactive zone. DLK-1 protein levels are elevated in rpm-1 mutants. RPM-1 can bind to, and stimulate ubiquitination of, DLK-1 in cultured cells. These data thus reveal an in vivo function for a previously uncharacterized p38 MAP kinase mediated signaling cascade in presynaptic development, and further suggest that activity of this MAP kinase cascade is negatively controlled by RPM-1 through regulation of protein stability. The subsynaptic localization of DLK-1 and RPM-1 implies that this regulation by RPM-1 may generate/maintain a “boundary” between cellular transporting domain and synaptic domain. FSN-1 shows physical and genetic interactions with an ALK-like receptor tyrosine kinase (Liao et al., 2004). It is yet to be investigated the functional connections between this ALK kinase and the DLK-1 pathway.
Synaptic assembly can be induced in vitro by multiple factors. Therefore, selecting synaptic partners is a crucial step in neural circuit formation (Scheiffele et al., 2000; Yamagata et al., 2003). Because target selection often occurs at the late stage of axon navigation, the mechanistic understanding of target recognition is complicated by the pleiotropic effect of genes that function in both axon guidance and target selection. One excellent system to examine this question is in the HSN neurons. HSN axons extend through half of the body, but form synapses specifically on two motor neurons, VC4 and VC5, at the mid-body near the vulva (Figure 2). Using a Punc-86SNB-1::YFP marker to label synaptic site of HSNs, it is shown that the vulval epithelium acts as a guidepost to instruct the HSN synapses (Shen and Bargmann, 2003). A screen for mutants displaying ectopic SNB-1::GFP in HSN neurons yielded two immunoglobulin superfamily genes syg-1 and syg-2 (Shen and Bargmann, 2003; Shen et al., 2004). SYG-1 is similar to the vertebrate NEPH1 and Drosophila IrrecC and Kirre/DUF, and SYG-2 is similar to vertebrate Nephrin and Drosophila Sticks and Stones (SNS) and Hibris (HIB). When expressed in Drosophila S2 cells, SYG-1 and SYG-2 proteins bind each other. syg-1 functions cell autonomously in HSNs. syg-2 is expressed in vulval epithelial cells. Functional SYG-1::GFP is localized to HSN synapses in wild type animals, but not in syg-2 mutants. These results demonstrate that the vulval epithelial functions as guidepost cells to instruct growth cone movements and that SYG-2 acts as a ligand for SYG-1 to initiate synapse formation. Not much is known about the molecules linking target recognition and assembly of synaptic components. SYD-1 protein contains PDZ, C2 and rhoGAP-like domains, and is localized at the presynaptic active zone. In syd-1 mutants different types of neurons show different phenotypes. In D type neurons, synaptic proteins are mislocalized and presynaptic terminals are formed in dendritic compartments in addition to their normal localizations (Hallam et al., 2002). In HSN neurons, SNB-1::GFP failed to accumulate at HSN synapses, rather retained in cell bodies. syd-1 may be involved in linking synaptic vesicular traffic and local cytoskeleton remodeling to membrane target recognition.
Many rudimentary aspects of synaptic assembly can happen independent of each other. Synapse formation depends on the recruitment of cell adhesion molecules (CAMs), which likely stabilize the cytoarchitecture in pre- and post- synaptic sites. In worm synapses, the endostatin-containing type XVIII collagen CLE-1 and Nidogen NID-1 display restricted localization pattern around synapses (Ackley et al., 2003). cle-1 mutants show enlarged SNB-1::GFP cluster, accompanied with reduced number of NMJ. nid-1 mutants have diffused SNB-1::GFP cluster. The function of NID-1 involves LAR RPTP, which interacts with SYD-2/Liprin to pattern presynaptic assembly (Ackley et al., 2005). An laminin alpha subunit, LAM-3, is expressed in nervous system (Huang et al., 2003). However, most of the mutations in laminins (epi-1, lam-3 and lam-1) cause early embryonic lethality (Huang et al., 2003; Zhu et al., 1999), precluding direct analysis of the role of laminins in synapse formation.
In vertebrate nervous system, synaptic activity plays a crucial role in shaping synaptic patterns. Not much is explored to address whether and how much activity plays a role in C. elegans. The cholinergic SAB neurons innervate head muscles, and show synapse sprouting and alterations in mutants with reduced cholinergic synaptic transmission. A source for this activity-dependence seems to involve a retrograde signal (Zhao and Nonet, 2000). UNC-122 is a conserved transmembrane protein containing collagen repeats and a cysteine-rich olfactomedin domain, and is localized to postsynaptic muscle arms (Loria et al., 2004). In unc-122 mutants NMJ development is morphological normal, the GABAergic DVB neuron displays axon sprouting due to impairment in synaptic transmission. It remains to be determined whether such axon and synapse spouting phenomena bear the same nature as those in other nervous system.
The identification of mutants from the primary visual screen using synaptic GFP reporter genes opens up the venue for further identification of synaptic components through genetic modifier screens and molecular binding partners studies. A major puzzle is how SV clustering and density formation is coordinated. In unc-104 animals, few synapses are detected with normal density (Hall and Hedgecock, 1991). Yet, most of the density proteins are localized largely normal by light microscope analysis (Hallam et al., 2002; Koushika et al., 2001). In syg-1 and syg-2 mutants, although HSN synapses are formed at incorrect locations, the ectopic synapses have similar, but small, configuration (Shen et al., 2004). Thus, SV clustering and density formation involve feedback mechanisms to which we know little about them. Understanding such mechanisms requires direct observation of synapse formation in real time. Necessary methodology that allows detection of synapse formation under low light and long period would greatly facilitate the studies of the dynamics of synaptic assembly. Ultrastructural analysis of synapses in young larvae would also be very informative.
Special thanks to M. Nonet, R. Weimer, J-L. Bessereau, D. Hall, for communicating unpublished results. I appreciate discussions and support from A. Chisholm and members of my laboratory.
Ackley, B.D., Kang, S.H., Crew, J.R., Suh, C., Jin, Y., and Kramer, J.M. (2003). The basement membrane components nidogen and type XVIII collagen regulate organization of neuromuscular junctions in Caenorhabditis elegans. J. Neurosci. 23, 3577–3587. Abstract
Ackley, B.D., Harrington, R.J., Hudson, M.L., Williams, L., Kenyon, C.J., Chisholm, A.D., Jin, Y. (2005). The two isoforms of the Caenorhabditis elegans leukocyte-common antigen related receptor tyrosine phosphatase PTP-3 function independently in axon guidance and synapse formation. J Neurosci. 25, 7517–7528. Abstract
Akert, K., Pfenninger, K., Sandri, C., and Moor, H. (1972). Freeze etching and cytochemistry of vesicles and membrane complexes in synapses of the central nervous system. In: Structure and Function of Synapses. G. D. Pappas, and D. P. Purpura, eds., Raven Press, New York, pp. 67–86.
Bodian, D. (1972). Synaptic diversity and characterization by electron microscopy. In: Structure and Function of Synapses. G. D. Pappas, and D. P. Purpura, eds., Raven Press, New York.
Burgess, R.W., Peterson, K.A., Johnson, M.J., Roix, J.J., Welsh, I.C., and O'Brien, T.P. (2004). Evidence for a conserved function in synapse formation reveals Phr1 as a candidate gene for respiratory failure in newborn mice. Mol. Cell Biol. 24, 1096–1105. Abstract
Byrd, D.T., Kawasaki, M., Walcoff, M., Hisamoto, N., Matsumoto, K., and Jin, Y. (2001). UNC-16, a JNK-signaling scaffold protein, regulates vesicle transport in C. elegans. Neuron 32, 787–800. Abstract
Cowan, W.M., Sudhof, T.C., and Stevens, C.F. (2001). Synapses. The Johns Hopkins University Press, Baltimore, Maryland.
Crump, J.G., Zhen, M., Jin, Y., and Bargmann, C.I. (2001). The SAD-1 kinase regulates presynaptic vesicle clustering and axon termination. Neuron 29, 115–129. Abstract
Deken, S.L., Vincent, R., Hadwiger, G., Liu, Q., Wang, Z.W., Nonet, M.L. (2005). Redundant localization mechanisms of RIM and ELKS in Caenorhabditis elegans. J Neurosci. 25, 5975–5983. Abstract
D'Souza, J., Hendricks, M., Le Guyader, S., Subburaju, S., Grunewald, B., Scholich, K., and Jesuthasan, S. (2005). Formation of the retinotectal projection requires Esrom, an ortholog of PAM (protein associated with Myc). Development 132, 247–256. Abstract Article
De Camilli, P., Haucke, V., Takai, K., and Mugnaini, E. (2001). The structure of synapses. In: Synapses, W.M. Cowan, T.C. Sudhof, and C.F. Stevens, eds., Baltimore, The Johns Hopkins University Press, pp. 89–134.
Goda, Y., and Davis, G.W. (2003). Mechanisms of synapse assembly and disassembly. Neuron 40, 243–264. Abstract
Greengard, P., Benfenati, F., and Valtorta, F. (1994). Synapsin I, an actin-binding protein regulating synaptic vesicle traffic in the nerve terminal. Adv. Second Messenger Phosphoprotein Res. 29, 31–45. Abstract
Hall, D.H., and Hedgecock, E.M. (1991). Kinesin-related gene unc-104 is required for axonal transport of synaptic vesicles in C. elegans. Cell 65, 837–847. Abstract
Hallam, S.J., Goncharov, A., McEwen, J., Baran, R., and Jin, Y. (2002). SYD-1, a presynaptic protein with PDZ, C2 and rhoGAP-like domains, specifies axon identity in C. elegans. Nat. Neurosci. 5, 1137–1146. Abstract Article
Hammarlund, M., Davis, W.S., and Jorgensen, E.M. (2000). Mutations in beta-spectrin disrupt axon outgrowth and sarcomere structure. J. Cell Biol. 149, 931–942. Abstract
Harrington, R.J., Gutch, M.J., Hengartner, M.O., Tonks, N.K., and Chisholm, A.D. (2002). The C. elegans LAR-like receptor tyrosine phosphatase PTP-3 and the VAB-1 Eph receptor tyrosine kinase have partly redundant functions in morphogenesis. Development 129, 2141–2153. Abstract
Harris, T.W., Hartwieg, E., Horvitz, H.R., and Jorgensen, E.M. (2000). Mutations in synaptojanin disrupt synaptic vesicle recycling. J. Cell Biol. 150, 589–600. Abstract
Huang, C.C., Hall, D.H., Hedgecock, E.M., Kao, G., Karantza, V., Vogel, B.E., Hutter, H., Chisholm, A.D., Yurchenco, P.D., and Wadsworth, W.G. (2003). Laminin alpha subunits and their role in C. elegans development. Development 130, 3343–3358. Abstract
Kishi, M., Pan, Y.A., Crump, J.G., and Sanes, J.R. (2005). Mammalian SAD kinases are required for neuronal polarization. Science 307, 929–932. Abstract Article
Ko, J., Na, M., Kim, S., Lee, J.R., and Kim, E. (2003). Interaction of the ERC family of RIM-binding proteins with the liprin-alpha family of multidomain proteins. J. Biol. Chem. 278, 42377–42385. Abstract Article
Koushika, S.P., Richmond, J.E., Hadwiger, G., Weimer, R.M., Jorgensen, E.M., and Nonet, M.L. (2001). A post-docking role for active zone protein Rim. Nat. Neurosci. 4, 997–1005. Abstract Article
Liao, E.H., Hung, W., Abrams, B., and Zhen, M. (2004). An SCF-like ubiquitin ligase complex that controls presynaptic differentiation. Nature 430, 345–350. Abstract Article
Loria, P.M., Hodgkin, J., and Hobert, O. (2004). A conserved postsynaptic transmembrane protein affecting neuromuscular signaling in Caenorhabditis elegans. J. Neurosci. 24, 2191–2201. Abstract Article
Murthy, V.N., and De Camilli, P. (2003). Cell biology of the presynaptic terminal. Ann. Rev. Neurosci. 26, 701–728. Abstract Article
Nakata, K., Abrams, B., Grill, B., Goncharov, A., Huang, X., Chisholm, A.D., and Jin, Y. (2005). Regulation of a DLK-1 and p38 MAP kinase pathway by the ubiquitin ligase RPM-1 is required for presynaptic development. Cell 120, 407–420. Abstract Article
Nguyen, M., Alfonso, A., Johnson, C.D., and Rand, J.B. (1995). Caenorhabditis elegans mutants resistant to inhibitors of acetylcholinesterase. Genetics 140, 527–535. Abstract
Nonet, M.L. (1999). Visualization of synaptic specializations in live C. elegans with synaptic vesicle protein-GFP fusions. J. Neurosci. Methods 89, 33–40. Abstract
Nonet, M.L., Holgado, A.M., Brewer, F., Serpe, C.J., Norbeck, B.A., Holleran, J., Wei, L., Hartwieg, E., Jorgensen, E.M., and Alfonso, A. (1999). UNC-11, a Caenorhabditis elegans AP180 homologue, regulates the size and protein composition of synaptic vesicles. Mol. Biol. Cell 10, 2343–2360. Abstract
Norman, K.R., and Moerman, D.G. (2002). Alpha spectrin is essential for morphogenesis and body wall muscle formation in Caenorhabditis elegans. J. Cell Biol. 157, 665–677. Abstract Article
O'Grady, P., Thai, T.C., and Saito, H. (1998). The laminin-nidogen complex is a ligand for a specific splice isoform of the transmembrane protein tyrosine phosphatase LAR. J. Cell Biol. 141, 1675–1684. Abstract
Ohtsuka, T., Takao-Rikitsu, E., Inoue, E., Inoue, M., Takeuchi, M., Matsubara, K., Deguchi-Tawarada, M., Satoh, K., Morimoto, K., Nakanishi, H., and Takai, Y. (2002). Cast: a novel protein of the cytomatrix at the active zone of synapses that forms a ternary complex with RIM1 and munc13-1. J. Cell Biol. 158, 577–590. Abstract Article
Ozaki, N., Shibasaki, T., Kashima, Y., Miki, T., Takahashi, K., Ueno, H., Sunaga, Y., Yano, H., Matsuura, Y., Iwanaga, T., et al. (2000). cAMP-GEFII is a direct target of cAMP in regulated exocytosis. Nat. Cell Biol. 2, 805–811. Abstract Article
Patel, N., Thierry-Mieg, D., and Mancillas, J.R. (1993). Cloning by insertional mutagenesis of a cDNA encoding Caenorhabditis elegans kinesin heavy chain. Proc. Natl. Acad. Sci. USA 90, 9181–9185. Abstract
Patton, B.L., Miner, J.H., Chiu, A.Y., and Sanes, J.R. (1997). Distribution and function of laminins in the neuromuscular system of developing, adult, and mutant mice. J. Cell Biol. 139, 1507–1521. Abstract
Richmond, J.E., Davis, W.S., and Jorgensen, E.M. (1999). UNC-13 is required for synaptic vesicle fusion in C. elegans. Nat. Neurosci. 2, 959–964. Abstract Article
Rongo, C., Whitfield, C.W., Rodal, A., Kim, S.K., and Kaplan, J.M. (1998). LIN-10 is a shared component of the polarized protein localization pathways in neurons and epithelia. Cell 94, 751–759. Abstract
Rostaing, P., Weimer, R.M., Jorgensen, E.M., Triller, A., and Bessereau, J.L. (2004). Preservation of immunoreactivity and fine structure of adult C. elegans tissues using high-pressure freezing. J. Histochem. Cytochem. 52, 1–12. Abstract
Saifee, O., Wei, L., and Nonet, M.L. (1998). The Caenorhabditis elegans unc-64 locus encodes a syntaxin that interacts genetically with synaptobrevin. Mol. Biol. Cell 9, 1235–1252. Abstract
Sakamoto, R., Byrd, D.T., Brown, H.M., Hisamoto, N., Matsumoto, K., and Jin, Y. (2004). The C. elegans UNC-14 RUN domain protein binds to the Kinesin-1/UNC-16 complex and regulates synaptic vesicle localization. Mol. Biol. Cell. Abstract Article
Schaefer, A.M., Hadwiger, G.D., and Nonet, M.L. (2000). rpm-1, a conserved neuronal gene that regulates targeting and synaptogenesis in C. elegans. Neuron 26, 345–356. Abstract
Scheiffele, P., Fan, J., Choih, J., Fetter, R., and Serafini, T. (2000). Neuroligin expressed in nonneuronal cells triggers presynaptic development in contacting axons. Cell 101, 657–669. Abstract
Schoch, S., Castillo, P.E., Jo, T., Mukherjee, K., Geppert, M., Wang, Y., Schmitz, F., Malenka, R.C., and Sudhof, T.C. (2002). RIM1alpha forms a protein scaffold for regulating neurotransmitter release at the active zone. Nature 415, 321–326. Abstract Article
Schuske, K.R., Richmond, J.E., Matthies, D.S., Davis, W.S., Runz, S., Rube, D.A., van der Bliek, A.M., and Jorgensen, E.M. (2003). Endophilin is required for synaptic vesicle endocytosis by localizing synaptojanin. Neuron 40, 749–762. Abstract
Serra-Pages, C., Medley, Q.G., Tang, M., Hart, A., and Streuli, M. (1998). Liprins, a family of LAR transmembrane protein-tyrosine phosphatase-interacting proteins. J. Biol. Chem. 273, 15611–15620. Abstract
Shen, K., and Bargmann, C.I. (2003). The immunoglobulin superfamily protein SYG-1 determines the location of specific synapses in C. elegans. Cell 112, 619–630. Abstract
Shen, K., Fetter, R.D., and Bargmann, C.I. (2004). Synaptic specificity is generated by the synaptic guidepost protein SYG-2 and its receptor, SYG-1. Cell 116, 869–881. Abstract
Signor, D., Wedaman, K.P., Rose, L.S., and Scholey, J.M. (1999). Two heteromeric kinesin complexes in chemosensory neurons and sensory cilia of Caenorhabditis elegans. Mol. Biol. Cell 10, 345–360. Abstract
Sorra, K.E., and Harris, K.M. (2000). Overview on the structure, composition, function, development, and plasticity of hippocampal dendritic spines. Hippocampus 10, 501–511. Abstract
Tam, T., Mathews, E., Snutch, T.P., and Schafer, W.R. (2000). Voltage-gated calcium channels direct neuronal migration in Caenorhabditis elegans. Dev. Biol. 226, 104–117. Abstract Article
Wan, H.I., DiAntonio, A., Fetter, R.D., Bergstrom, K., Strauss, R., and Goodman, C.S. (2000). Highwire regulates synaptic growth in Drosophila. Neuron 26, 313–329. Abstract
Wang, Y., Liu, X., Biederer, T., and Sudhof, T.C. (2002). A family of RIM-binding proteins regulated by alternative splicing: implications for the genesis of synaptic active zones. Proc. Natl. Acad. Sci. USA 99, 14464–14469. Abstract Article
Wang, Y., Okamoto, M., Schmitz, F., Hofmann, K., and Sudhof, T.C. (1997). Rim is a putative Rab3 effector in regulating synaptic-vesicle fusion. Nature 388, 593–598. Abstract Article
Weimer, R.M., Richmond, J.E., Davis, W.S., Hadwiger, G., Nonet, M.L., and Jorgensen, E.M. (2003). Defects in synaptic vesicle docking in unc-18 mutants. Nat. Neurosci. 6, 1023–1030. Abstract Article
White, J.G., Southgate, E., Thomson, J.N., and Brenner, S. (1986). The structure of the nervous system of the nematode C. elegans. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 314B, 1–340.
Wyszynski, M., Kim, E., Dunah, A.W., Passafaro, M., Valtschanoff, J.G., Serra-Pages, C., Streuli, M., Weinberg, R.J., and Sheng, M. (2002). Interaction between GRIP and liprin-alpha/SYD2 is required for AMPA receptor targeting. Neuron 34, 39–52. Abstract
Yamagata, M., Sanes, J.R., and Weiner, J.A. (2003). Synaptic adhesion molecules. Curr. Opin. Cell Biol. 15, 621–632. Abstract
Yeh, E., Kawano, T., Weimer, R.M., Bessereau, J.L., and Zhen, M. (2005). Identification of genes involved in synaptogenesis using a fluorescent active zone marker in Caenorhabditis elegans. J Neurosci. 25, 3833–3841. Abstract
Zhai, R.G., and Bellen, H.J. (2004). The architecture of the active zone in the presynaptic nerve terminal. Physiology (Bethesda) 19, 262–270. Abstract Article
Zhao, H., and Nonet, M.L. (2000). A retrograde signal is involved in activity-dependent remodeling at a C. elegans neuromuscular junction. Development 127, 1253–1266. Abstract
Zhen, M., Huang, X., Bamber, B., and Jin, Y. (2000). Regulation of presynaptic terminal organization by C. elegans RPM-1, a putative guanine nucleotide exchanger with a RING-H2 finger domain. Neuron 26, 331–343. Abstract
Zhen, M., and Jin, Y. (1999). The liprin protein SYD-2 regulates the differentiation of presynaptic termini in C. elegans. Nature 401, 371–375. Abstract Article
*Edited by Erik M. Jorgensen and Joshua M. Kaplan. Last revised December 9, 2005. Published December 23, 2005. This chapter should be cited as: Jin, Y. Synaptogenesis (December 23, 2005), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.44.1, http://www.wormbook.org.
Copyright: © 2005 Yishi Jin, This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
§To whom correspondence should be addressed. E-mail: jin@biology.ucsc.edu
 All journal content, except where otherwise noted, is licensed under a Creative Commons Attribution License.
All journal content, except where otherwise noted, is licensed under a Creative Commons Attribution License.