Embryo series courtesy of Einhard Schierenberg
Embryo series courtesy of Einhard SchierenbergTable of Contents
Abstract
Appropriate regulation of mRNA transcription is central to the differentiation and functions of eukaryotic cells, and to the development of complex organisms. mRNAs are synthesized by the coordinated action of a set of general transcription and mRNA modification factors. These factors and the fundamental mechanisms involved in transcription are conserved among eukaryotes, including C. elegans. Recent studies in various systems have revealed that this apparatus is not controlled through a simple on/off “switch” at the promoter, and that the factors and mechanisms involved in transcription are instead subject to regulation at a surprising number of different levels. In this chapter we will discuss examples in which regulation involving the general mRNA transcription apparatus or other transcription co-factors plays a central role in C. elegans development, and in which C. elegans studies have provided new insights into eukaryotic transcription mechanisms. Together, these studies have shown that regulatory mechanisms that involve the general Pol II machinery are a central participant in many aspects of C. elegans biology.
In this first section we will briefly describe the processes through which eukaryotic mRNAs are synthesized by RNA Polymerase (Pol) II. In general, the findings discussed in this section have been obtained in cellular or in vitro studies of S. cerevisiae, Drosophila embryos, or cultured human cells, but we will also refer to aspects of C. elegans biology where indicated. In subsequent sections of the chapter, we will discuss how analyses of these fundamental mRNA transcription mechanisms or factors have provided important insights into C. elegans biology, or vice versa. We will not discuss RNA Pol I (ribosomal RNA genes) or Pol III (small nuclear and transfer RNA genes), which have been studied very little in C. elegans.
Eukaryotic mRNAs are synthesized by Pol II through an intricate multistep process (Lemon and Tjian, 2000; Orphanides and Reinberg, 2002). mRNA transcription initiates at a fixed start site that is located adjacent to the promoter (Figure 1A). Transcription then must be maintained during an elongation phase, in which Pol II progresses more distally along the gene. Remarkably, these events are mechanistically coupled to the series of steps that process the transcript into a mature mRNA. Although very few biochemical studies of C. elegans transcription complexes have been performed, it seems almost certain that these basic mRNA transcription mechanisms have been conserved in C. elegans. Firstly, most of the components of the C. elegans general transcription machinery have been identified at the DNA sequence level (Table 1). In addition, as described in the text, in vivo studies of C. elegans transcription machinery components have generally yielded results consistent with mechanistic functions that were defined in other systems.
|
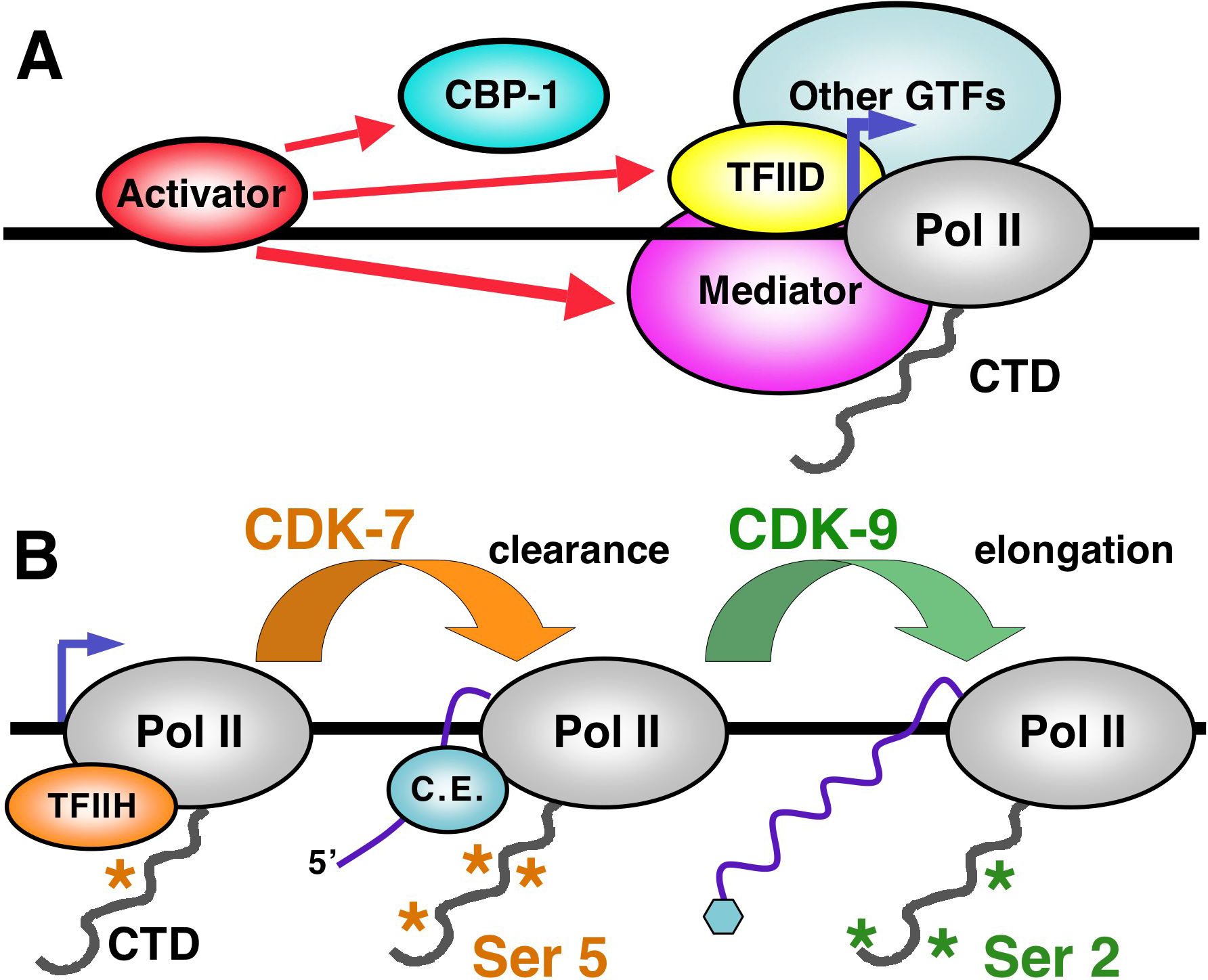
Figure 1. Transcription regulatory complexes. (A) Partial view of the Pol II pre-initiation complex (Orphanides and Reinberg, 2002), subunits of which include Mediator and a set of GTFs (TFIIA, TFIIB, TFIID, TFIIE, TFIIH, and others; Table 1). Most pre-initiation complex factors, including Pol II, consist of multiple subunits. Interactions between pre-initiation complex subunits and core promoter elements localize formation of the pre-initiation complex, which in turn sets the transcription start site (arrow). Pre-initiation complex formation is driven by recruitment of pre-initiation complex components through interactions with DNA-binding activators (see Transcription regulation), which at some genes also recruit chromatin-modifying co-factors such as the HAT CBP-1 (see text). (B) The Pol II CTD phosphorylation cycle (Bentley, 2005; Buratowski, 2005; Orphanides and Reinberg, 2002). After Pol II has been recruited into a pre-initiation complex, the CTD repeat is phosphorylated on Ser 5 by the CDK-7 subunit of the GTF TFIIH. This phosphorylation is required for Pol II to transcribe beyond the immediate promoter region (clearance), and for recruitment of the mRNA capping enzyme. Subsequently, phosphorylation of CTD Ser 2 by CDK-9 facilitates elongation and is required for mRNA termination, cleavage, and processing (not shown). CDK-9 is a subunit of the GTF P-TEFb (positive transcription elongation factor b, Table 1).
The first step in eukaryotic mRNA transcription is assembly of a pre-initiation complex (PIC) that consists of Pol II, a set of conserved general transcription factors (GTFs), and Mediator, a large multiprotein complex that communicates directly with many gene-specific regulators (Figure 1A; Green, 2005; Kornberg, 2005; Lemon and Tjian, 2000; Orphanides and Reinberg, 2002). As described in Section 5, genetic studies of C. elegans Mediator subunits have synergized beautifully with biochemical and functional experiments to link these subunits to specific transcription pathways and biological processes. PIC assembly is directed to the start site by “core” promoter sequences that are recognized primarily by subunits of the GTF TFIID, including the TATA-binding protein (TBP), or by related protein complexes (Section 4; Muller and Tora, 2004). This process is driven by upstream gene-specific activators that bind to regulatory sequences within promoters or distal enhancer elements, and recruit PIC components to the promoter through direct interactions (Figure 1A). The complexity involved in PIC assembly makes it possible to integrate many regulatory signals at the promoter, making it possible for an individual gene to be responsive to multiple independent cues.
Recruitment of the PIC by gene-specific activators is the most fundamental mechanism through which transcription is induced, but PIC assembly is also influenced profoundly by the positioning or modification of histones, the basic building block of chromatin. The configuration of chromatin at the promoter region is in turn affected through recruitment of chromatin modification or remodeling factors by gene-specific activators or repressors. Transcription regulation at the chromatin level is an increasingly complex subject that is largely beyond the scope of this chapter, and has been addressed partially in other chapters (see Germline chromatin, Specification of the germ line), but we will briefly describe some examples in which interplays among chromatin remodelers have important functions in C. elegans biology (Section 6).
In C. elegans numerous promoters that regulate expression of individual genes have been identified and mapped (see Transcription regulation), but little is known about the core promoter elements that direct the Pol II machinery to specific transcription start sites. The basic PIC apparatus that recognizes core promoters is present in C. elegans, but approximately 70% of C. elegans genes are trans-spliced to one of a small number of common leader sequences (see Trans-splicing and operons), making it almost impossible to identify their transcription start sites precisely. Some predicted C. elegans core promoters include apparent TATA elements but others do not (see Transcription regulation), as is true in other organisms, but the counterparts to other core promoter elements identified in other systems (Smale and Kadonaga, 2003) have not been studied in C. elegans. At the same time, known core promoter recognition factors are critical for transcription in C. elegans (Section 4), indicating that regulatory principles identified elsewhere are likely to apply. C. elegans upstream regulatory regions are more compact than those of higher eukaryotes (usually <2kb, see Transcription regulation), and generally do not rely on distant “enhancer” elements, but similarly include clusters of sites for gene-specific regulators. Our understanding of the architecture of gene-specific C. elegans regulatory regions will continue to increase with expansion of genomics resources for characterizing these elements and identifying proteins that bind to them (Deplancke et al., 2006; Dupuy et al., 2004).
Elegant mechanistic analyses in other systems have shown that while transcription can begin after assembly of the PIC is complete, subsequent transcription steps depend upon phosphorylation of the Pol II large subunit C-terminal domain (CTD) (Figure 1B; Bentley, 2005; Buratowski, 2005; Meinhart et al., 2005). The CTD consists of tandem repeats (38 in C. elegans, 52 in humans) that are based upon the sequence YSPTSPS. Pol II is recruited with the CTD in an unphosphorylated form, then at the promoter the CTD repeat is phosphorylated on Ser 5 by cyclin-dependent kinase (CDK)-7. This phosphorylation allows Pol II to move away from the promoter region, and results in recruitment of the mRNA capping enzyme (C. E., Figure 1B). The CTD is then phosphorylated on Ser 2 by CDK-9 (Figure 1B). Ser 2 phosphorylation promotes transcription elongation, and allows mRNA termination and processing factors to be recruited directly to the CTD. One of the most interesting concepts to emerge from the transcription field during the past decade is that the CTD acts as a molecular integrator that physically couples transcription to downstream nuclear events that include mRNA processing, termination, and even nuclear export, and that the cycle of CTD phosphorylation is critical for orchestrating these events. Dephosphorylation of the CTD may occur in competition with the action of CTD kinases (not shown), and is required for Pol II to be recycled for future transcription. As is described below, a number of studies have investigated the regulation of C. elegans embryonic transcription in part by monitoring CTD phosphorylation (Sections 3 and 4).
Because of its considerable advantages for genetic, phenotypic, and cell biological studies, most analyses of transcription mechanisms in C. elegans have involved intact embryos or animals. One of the most important contributions of C. elegans to the transcription field is that mutations in PIC genes have been recovered in genetic screens, and have linked regulation that involves these factors to specific biological processes (Section 5 and 6). Another advantage of C. elegans is that during early embryonic stages both maternal and zygotic gene expression can be disrupted by RNA interference (RNAi), and viability can be sustained by maternal mRNAs in the absence of transcription. For example, without the Pol II catalytic large subunit AMA-1 embryonic cells continue dividing to form a cluster of approximately 100 undifferentiated cells (Powell-Coffman et al., 1996), even though transcription of zygotic genes normally begins much earlier (Section 3). It has therefore been possible in the early embryo to study functions of GTFs that are otherwise essential for cell viability (Section 4).
A variety of techniques have been employed to assess mRNA production or occurrence of transcription steps in living or fixed C. elegans. One of the earliest approaches to be adopted was measurement or visualization of labeled UTP incorporation (Edgar et al., 1994; Gibert et al., 1984; Schisa et al., 2001). This technique provides a rough but direct indicator of mRNA production over time, and has been used to assess when high-level transcription begins during embryogenesis and germ cell development, and to track mRNA localization. A major advance for studying C. elegans gene regulation was the development of techniques for visualizing the presence of individual mRNAs by in situ hybridization (Seydoux and Fire, 1994). The most widely used means of assaying gene expression in C. elegans is production of transgenic reporter strains in which a coding region or predicted promoter of interest is fused to green fluorescent protein (GFP), or another marker that can be detected in living animals. This approach, and its advantages and potential pitfalls, are described in detail in the Transcription regulation chapter. An indirect but valuable technique for studying transcription has been to perform immunostaining or western blotting with well-characterized antibodies that recognize Ser 5- or Ser 2-phosphorylated forms of the CTD repeat (Bentley, 2005; Buratowski, 2005), and are refered to here as α-PSer5 and α-PSer2. These assays provide approximate indicators of the occurrence of steps associated with the initiation and elongation phases of transcription, respectively (Section 4).
In C. elegans in vitro studies of transcription mechanisms have been performed relatively infrequently, although most biochemical techniques through which transcription has been investigated in other systems are feasible. One of the earliest investigations of transcription in C. elegans involved a series of parallel biochemical and genetic studies of RNA polymerase activity. RNA Pol I, Pol II, and Pol III were partially purified from C. elegans, and shown to parallel the corresponding polymerases from other organisms with respect to their relative sensitivies to the mushroom toxin α-amanitin (Sanford et al., 1983; Sanford et al., 1985). At the same time, mutagenesis screens for increased resistance to α-amanitin identified multiple mutations in the ama-1 gene that resulted in production of α-amanitin-resistant Pol II (Rogalski et al., 1988; Sanford et al., 1983). ama-1 was then shown to encode the Pol II large subunit based upon its similarity to the corresponding Drosophila gene (Bird and Riddle, 1989). Subsequently, a C. elegans Pol II in vitro transcription system was developed in which transcription is induced appropriately and specifically by promoter-bound activators (Lichtsteiner and Tjian, 1993; Lichtsteiner and Tjian, 1995). This system has not become widely used however, most likely because other organisms are still more advantageous for mechanistic studies since their PIC subunits and other basic transcription machinery components have been characterized more extensively at the biochemical level.
More recently, C. elegans protocols have been developed for studying transcription factor functions through chromatin immunoprecipitation (ChIP; Chu et al., 2002; Oh et al., 2006; Whetstine et al., 2005), a technique that assays for direct binding of proteins to specific DNA sequences within cells, and is used widely in yeast and mammalian cell studies. In ChIP, transcription factors and bound DNA are crosslinked together in intact cells in the context of chromatin, which is sheared after cell lysis. DNA that is bound by the protein of interest is then identified by co-immunoprecipitation and PCR. With this technique, it is possible to test whether a transcription factor of interest binds directly to specific loci in the genome. A very exciting recent study showed that in C. elegans ChIP can be used to identify novel sequences bound by a transcription factor, in this case the regulator of Dauer development and longevity DAF-16 (Oh et al., 2006). As a greater number of antibodies and transgenic tools become available, in C. elegans it will be possible to use ChIP not only for an expanded range of investigative experiments, but potentially to identify new targets of any transcription factor of interest.
Across diverse species, mRNA transcription is globally shut down during oogenesis then reactivated early during embryonic development (Davidson, 1986). In C. elegans, this shutdown occurs as oocytes enter the diakinesis stage of meiotic prophase and begin to undergo maturation, as indicated by dramatic reductions in labeled UTP incorporation and α-PSer5 staining (Kelly et al., 2002; Schisa et al., 2001). Transcriptional silence is then maintained until the 4-cell stage of embryonic development (Seydoux and Dunn, 1997). It is not understood in any species how this transcriptional silencing is established, or is later relieved in the embryo. One rationale for having this period of transcription silence is that it may be advantageous for the earliest stages of embryonic development to be orchestrated entirely through post-translational regulation of "stored" maternal gene products, a strategy that may provide the most rapid and versatile means of modulating gene expression spatially and temporally during this critical period. It is also possible that this transcriptional silencing facilitates the transition between maternal and embryonic transcription programs that are very different from each other (Baugh et al., 2003).
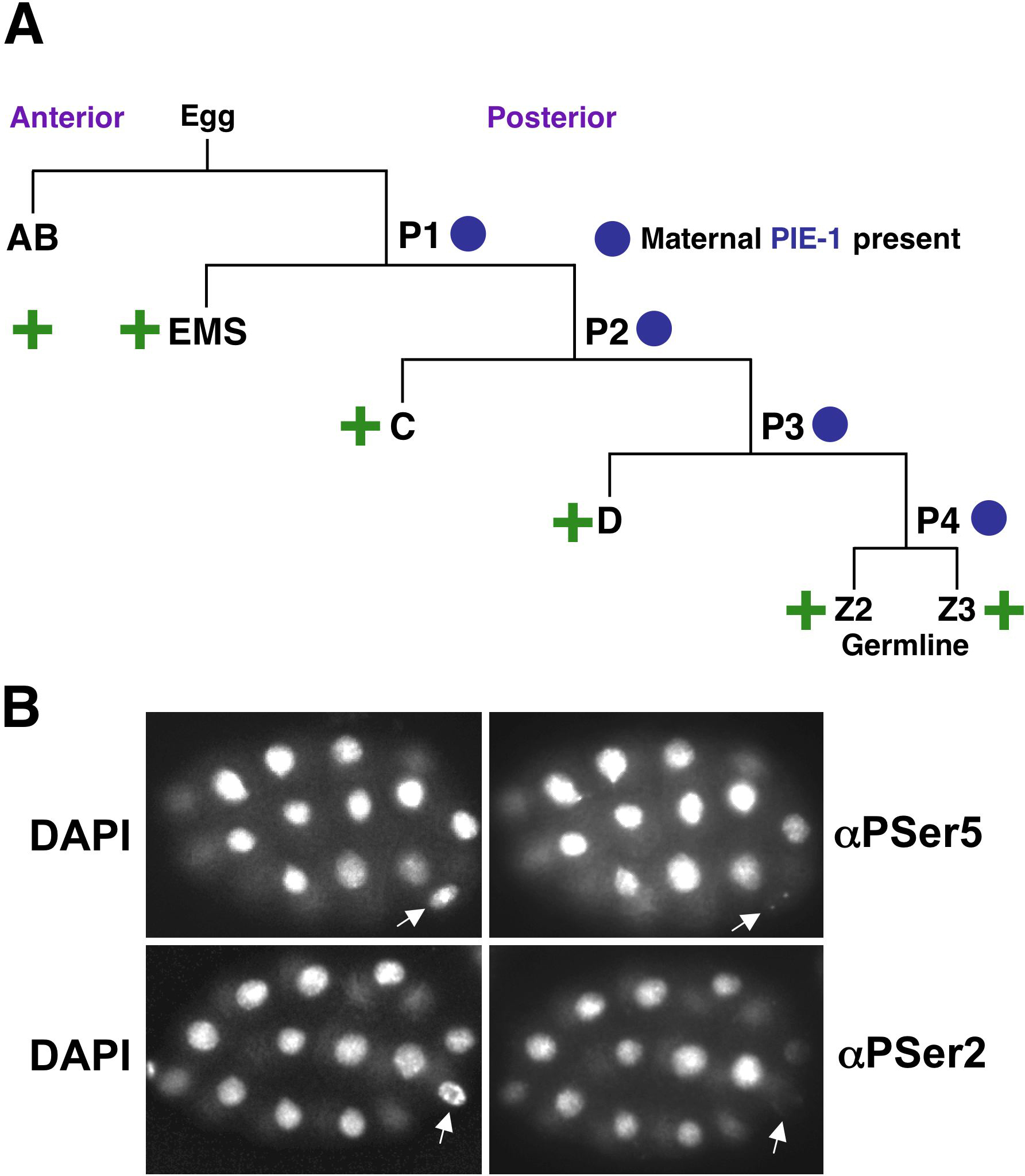
In the developing embryo, the onset of new transcription is delayed in the germline relative to somatic blastomeres. Analyses of individual genes and phospho-CTD antibody staining indicate that while transcription begins at the 4-cell stage in somatic lineages, transcriptional silence is maintained in embryonic germ cell precursors until they have been segregated from somatic lineages, after the 100-cell stage (Figure 2A, B; Mello et al., 1992; Seydoux and Dunn, 1997). This delay depends upon the germline-specific protein PIE-1, without which germ cell precursors become transcriptionally active in parallel to their zygotic counterparts, and follow somatic differentiation programs (Seydoux et al., 1996).
|
Figure 2. Delayed transcription onset in the embryonic germline. (A) Embryonic founder cell diagram showing that the beginning of new transcription (green plus sign) is delayed in the germline until after formation of Z2 and Z3, and disappearance of PIE-1 (Seydoux and Dunn, 1997). (B) Distinct CTD phosphorylation patterns in embryonic somatic and germline lineage cells (Seydoux and Dunn, 1997). These 28-cell embryos are stained with DAPI (left) and the indicated phospho-CTD antibodies (right). Note the characteristic pair of αPSer5-staining dots and lack of αPSer-2 staining in the germline blastomere (arrows). Posterior is to the right.
PIE-1 is a zinc finger protein of an RNA-binding class (Seydoux et al., 1996), making it a particularly intriguing question how it might repress transcription. PIE-1 regions potently block transcription when they are tethered to promoters in mammalian cells (Batchelder et al., 1999), and its silencing of embryonic germline transcription depends upon its presence in nuclei (Tenenhaus et al., 2001). One PIE-1 region interferes with CDK-9 function and inhibits elongation, dependent upon a motif that resembles an unphosphorylatable CTD repeat and contributes to PIE-1 function in vivo (Batchelder et al., 1999; Zhang et al., 2003). Interestingly, transcriptionally silent embryonic germ cell precursors lack detectable α-PSer2 staining, but contain two α-PSer5 foci that depend upon presence of transcription initiation factors (Figure 2B; Seydoux and Dunn, 1997; Shim et al., 2002a; Walker et al., 2001). Taken together, this evidence suggests that PIE-1 may block transcription directly at promoters, after some initiation steps have been completed. Consistent with this model, markers associated with an "active" chromatin configuration are comparably present in early embryonic germline and somatic precursors, suggesting that this silencing of germline transcription is not mediated through global chromatin effects (Schaner et al., 2003; see Germline chromatin). It is not known how PIE-1 is brought to individual genes, but a speculative possibility is that its zinc fingers might bind nascent mRNAs. In addition to this transcriptional silencing role PIE-1 has additional functions that involve translational regulation, and chromatin modification at germline-specific genes (Tenenhaus et al., 2001; Unhavaithaya et al., 2002; see Specification of the germ line). Genetic and site-directed mutagenesis evidence indicates that each of these latter PIE-1 functions are mediated through different mechanisms from its transcriptional repressor activity however, and therefore that PIE-1 is involved in multiple distinct germline processes.
It may be advantageous for transcription of embryonic genes to be inhibited at a post-initiation step in germline precursors, because this strategy would presumably allow the transcription block to be reversed very quickly. When embryonic germ cell precursors divide, evidence of new transcription is seen rapidly in each somatic daughter (Seydoux and Dunn, 1997), suggesting that some genes may have been poised for immediate expression. A PIE-1 ortholog has not been described outside of C. elegans, but in Drosophila embryonic germ cell precursors are also transcriptionally silent and similarly contain foci of Ser 5- but not Ser 2-phosphorylated Pol II (Seydoux and Dunn, 1997). Post-initiation inhibition of some transcription therefore may be a feature of germ cell precursors in various organisms.
As indicated in Section 2, the functions of even essential transcription factors can be investigated in the early C. elegans embryo because of the presence of maternal gene products. For example, conditional inactivation of the CTD kinase CDK-7 (Figure 1B) essentially abolishes transcription and CTD phosphorylation in living embryonic cells, as predicted by biochemical studies (Wallenfang and Seydoux, 2002). By showing that most of the Ser 5 phosphorylation detected in early embryos is cdk-7-dependent, this experiment indicated that this CTD phosphorylation is associated with transcription activity and does not derive from other kinases that can phosphorylate the CTD, but are not integral to the transcription process (Meinhart et al., 2005). Another question that has been addressed is the importance of phosphorylation of CTD Ser 2 by the PIE-1 target CDK-9 (Figure 1B). In vitro and cell culture studies had identified CDK-9 as an enhancer of elongation, particularly during HIV transcription, and had showed that it preferentially phosphorylates CTD Ser 2 (Price, 2000). Ser 2 can be phosphorylated by other kinases however, and in yeast the CDK-9 counterpart is not essential for viability, raising the question of whether CDK-9 is needed for most metazoan transcription events. cdk-9(RNAi) embryos are characterized by robust Ser 5 phosphorylation but undetectable Ser 2 phosphorylation, and an apparent absence of new gene expression, indicating that CDK-9 is the major Ser 2 kinase and is needed for most if not all early embryonic transcription (Shim et al., 2002b). RNAi knockdown of the C. elegans mRNA capping enzyme resulted in a similar phenotype, supporting the idea that this mRNA processing event represents an intermediate step between Ser 5 phosphorylation, and Ser 2 phosphorylation and elongation (Takagi et al., 2003; Figure 1B).
Other embryo studies have studied the core promoter recognition factor TFIID (Figure 1A), which includes TBP and a set of TFIID-associated factors (TAFIIs; Table 1; Tora, 2002). Many TAFIIs are also found in alternative complexes that bind mRNA start sites (Tora, 2002). While TBP is generally required for mRNA transcription, most TAFIIs have specialized functions that involve interactions with core promoter sequences or other transcription factors. The extent to which individual TAFIIs are required for mRNA transcription, as indicated by genetic studies in yeast, has remained controversial (see (Walker et al., 2004)). In C. elegans a TFIID-specific TAFII (TAF-1) is required at most embryonic genes (Walker et al., 2004), in contrast to evidence from yeast that an alternative core promoter complex can substitute at many genes (Huisinga and Pugh, 2004). This predicts that core promoter elements that interact with TAF-1 may be critical at most C. elegans embryonic genes. One group of C. elegans TAFIIs (TAF-5, -9, -10) is required broadly for embryonic transcription but is dispensible at most genes that are specific to metazoa, suggesting that the latter genes have diverged in their regulation (Walker and Blackwell, 2003; Walker et al., 2001). In each case, after C. elegans core promoter elements have been better characterized, it should be possible to understand why these TAFIIs are required for expression of some genes but not others. Surprisingly, TAF-4 is needed for essentially all early embryonic transcription, making it particularly interesting to elucidate its mechanistic function (Walker et al., 2001). Interestingly, the α-PSer5 and α-PSer2 staining patterns of taf-4(RNAi) somatic embryonic cells appear identical to those normally seen in embryonic germ cell precursors (see Figure 2B; Walker et al., 2001), suggesting parallels between how transcription is blocked by PIE-1 (Section 3) and lack of TAF-4. A speculative model is that in each case transcription has been stalled prior to Ser 2 phosphorylation, and that the Ser 5-phosphorylated Pol II that is generated is recycled within the two characteristic foci (Figure 2B).
Given that TFIID and TBP are broadly essential for mRNA transcription, it is intriguing that in C. elegans most early embryonic transcription requires the TBP-related protein TLF-1 (Dantonel et al., 2000; Kaltenbach et al., 2000). Studies in other organisms indicate that TLF does not associate with TAFIIs, or form a TFIID-like complex (Chong et al., 2005; Hochheimer et al., 2002). TLF is important for some early embryonic transcription in Xenopus and Zebrafish but not in mice (Martianov et al., 2001; Muller et al., 2001; Veenstra et al., 2000), in which it is required specifically for chromatin reorganization during spermatogenesis (Martianov et al., 2002). One possible model is that TLF might influence chromatin organization during early stages in the C. elegans embryo, when gene expression patterns are changing rapidly (Baugh et al., 2003).
Among the various components of the Pol II PIC (Figure 1A), Mediator appears to be the most important integrator of regulatory signals from upstream activators that define specific patterns of gene expression (Green, 2005; Kornberg, 2005). Biochemical and genetic studies indicate that the basic features of Mediator are conserved across species, and that Mediator projects many regulatory surfaces that interact with gene-specific activators or repressors (Bjorklund and Gustafsson, 2005; Green, 2005; Malik and Roeder, 2005; Wang et al., 2005). C. elegans orthologs of 24 of the 33 mammalian Mediator subunits have been identified, primarily through sequence similarities (Table 1; Bourbon et al., 2004; Kwon et al., 1999; Yoda et al., 2005). The C. elegans Mediator complex(es) have not been thoroughly characterized biochemically, but some C. elegans Mediator proteins have been shown to co-fractionate or co-immunoprecipitate with each other (Kwon et al., 1999). Mediator subunits have recently been designated as MED proteins in other species, and MDT (MeDiaTor) proteins in C. elegans (Bourbon et al., 2004). Here I refer to C. elegans Mediator subunits primarily by their original designations, referencing the unified nomenclature in parentheses. Some C. elegans Mediator subunits have pleiotropic functions (Kwon and Lee, 2001; Kwon et al., 1999). For example, RGR-1 (MDT-14) appears to be needed for essentially all early embryonic transcription (Shim et al., 2002a). Significantly however, mutations recovered in genetic screens and protein-protein interaction studies have implicated other Mediator subunits as being central for particular conserved transcription pathways.
Studies of C. elegans MED23 represent a particularly elegant synergy of genetic and biochemical approaches. In C. elegans MED23 was initially identified as the novel protein SUR-2 (MDT-23), through a screen for the Suppression of the Ras gain-of-function allele let-60(n1046) (Singh and Han, 1995). Genetic evidence indicated that sur-2 functions downstream of the Ras/Mitogen Activated Protein Kinase (MAPK) signaling pathway that induces the vulval cell fate, and may act at the level of the ETS transcription factor LIN-1, which is regulated by this signaling pathway. The molecular function of SUR-2 (MDT-23) remained a mystery however, until this protein was identified in a mammalian biochemical study as a Mediator component (Boyer et al., 1999). Mammalian SUR-2 was then found to be a direct target through which the adenovirus protein E1A recruits Mediator in order to activate transcription of adenovirus genes. C. elegans genetics and mammalian biochemistry thus together suggested an exciting model, that SUR-2 (MDT-23) might be a specific conduit through which Ras/MAPK-responsive transcription factors communicate with the general transcription machinery. Supporting this idea, mammalian MED23 was found to be required for MAPK-induced transcription, and to be targeted directly by a mammalian Ras/MAPK-activated ETS transcription factor, but not by most other activators (Boyer et al., 1999; Stevens et al., 2002).
The complexity and specificity of Mediator functions is illustrated further by analyses of C. elegans DPY-22 (MDT-12) and LET-19 (MDT-13). Structural analyses in other systems have shown that these proteins are present in a Mediator submodule that is thought to have mostly inhibitory functions (Bjorklund and Gustafsson, 2005; Malik and Roeder, 2005). Accordingly, genetic studies indicate that DPY-22 (MDT-12) acts antagonistically to Ras signaling during vulval development (Moghal and Sternberg, 2003), and that DPY-22 (MDT-12) and LET-19 (MDT-13) inhibit Wnt-induced gene expression in multiple contexts, supporting the idea that they function together (Wang et al., 2004; Yoda et al., 2005; Zhang and Emmons, 2000). The C. elegans MED1 ortholog sop-3 (mdt-1.1) has similar genetic properties (Zhang and Emmons, 2001). This suggests that MED1 may interact functionally with MED12 and MED13 even though it is not in the same Mediator submodule, or may communicate directly with the same regulatory factors. A strong RNAi knockdown of LET-19 (MDT-13) prevents expression of some early embryonic genes however, suggesting that this Mediator subunit also has some positive functions (Wang et al., 2004). By identifying specific transcription pathways in which DPY-22 (MDT-12) and LET-19 (MDT-13) are important, these studies have made it of particular interest to determine whether the corresponding transcription activators, co-factors, or repressors target these Mediator subunits directly, as is the case for SUR-2 (MDT-23).
Two exciting recent studies have further demonstrated the importance of Mediator for coordinating regulation of gene networks, and have shown the value of C. elegans for uncovering regulatory mechanisms that are of direct relevance to human health. The C. elegans Mediator subunit MDT-15 (MED15 in mammals) has been identified as a direct target of two transcription factors, the nuclear receptor NHR-49 and the sterol regulatory element binding protein (SREBP) (Taubert et al., 2006; Yang et al., 2006). Two-hybrid screening indicated that MDT-15 binds specifically to NHR-49, which functionally resembles human Peroxisome Proliferator-Activated Receptor-α, a regulator of lipid metabolism that is an important target of therapy in diabetes (Taubert et al., 2006). It was then determined that mdt-15 is required for expression of NHR-49-dependent genes, along with many other lipid metabolism genes. Similar results were obtained in a study of the transcription factor SREBP, a key regulator of cholesterol and lipid homeostasis (Yang et al., 2006). It was shown that MDT-15/MED15 interacts directly and specifically with SREBP, and is required for SREBP-driven transcription in human cells and C. elegans. In C. elegans, RNAi knockdown of either mdt-15 or sbp-1 (SREBP) perturbed fat storage, and resulted in alterations in the levels of specific lipids that were consistent with the accompanying abnormalities in lipid metabolism gene expression (Taubert et al., 2006; Yang et al., 2006). Remarkably, phenotypes associated with mdt-15 RNAi (decreased lifespan and fertility, locomotion impairment, abnormal fat storage) were then partially rescued by dietary supplementation with specific fatty acids. Together, these studies showed that MDT-15/MED15 integrates specific transcription pathways that regulate lipid and cholesterol homeostasis and that this MED15 function is conserved, a finding that may lead to a more comprehensive understanding of how lipid metabolism is regulated coordinately in humans. Through its interactions with SREBP and nuclear receptors, MED15 may therefore be involved in metabolic abnormalities that include obesity, atherosclerosis, diabetes, and metabolic syndrome.
As recently as fifteen years ago, it was thought that they key to understanding transcription regulation lay almost entirely in understanding how activators and repressors acted on the general transcription machinery, but since then it has become apparent that chromatin is a central mechanistic player in the regulation of many if not most eukaryotic genes. The field of chromatin regulation in C. elegans has expanded rapidly during recent years, in part through reverse genetic studies of the functions of specific chromatin regulators, but also because protein complexes that modify chromatin structure have appeared in mutagenesis or RNAi screens that involve specific biological processes. In C. elegans chromatin modifications are needed to establish some broad transcription programs, particularly with respect to discriminating between germline and somatic gene expression patterns (see Germline chromatin; Specification of the germ line). C. elegans chromatin modification complexes have also been implicated in regulation of a variety of specific biological functions or gene networks, however. This field is now much too broad to be covered comprehensively within the context of this chapter, but here we will describe some specific examples of how interactions among chromatin modifiers play critical roles in C. elegans transcription regulation and development.
The transcription regulators CBP-1 and HDA-1 provide an instructive example of how interactions among chromatin modification factors are important for specific biological processes during the life cycle. Acetylation and deacetylation of histone N-terminal tails is usually associated with increased or decreased transcription, respectively (Orphanides and Reinberg, 2002). CBP-1 is a conserved histone acetyl transferase (HAT) that in mammals often responds to extracellular signals, and is recruited to genes by upstream activators (Figure 1A; Chan and La Thangue, 2001). In the C. elegans embryo, CBP-1 is required for differentiation of essentially all cell types except neurons (Shi and Mello, 1998). Only a minority proportion of embryonic genes depend upon cbp-1 for their expression however (Walker et al., 2004), presumably because these genes are otherwise repressed through specific chromatin modifications that are counterracted by CBP-1. For example, the embryonic endoderm specification gene end-1 requires CBP-1 in order to overcome repression by HDA-1, a histone deacetylase that is recruited by gene-specific repressors (Calvo et al., 2001). Inhibition of hda-1 expression by RNAi allows end-1 to be expressed independently of CBP-1, and endodermal cells to be formed independently of either CBP-1 or SKN-1, the transcription factor that initiates endoderm specification in the embryo (Calvo et al., 2001; Shi and Mello, 1998). Like CBP-1, HDA-1 appears to regulate only a limited number of embryonic genes, but usually as a repressor (Whetstine et al., 2005). Microarray and ChIP experiments have shown that HDA-1 directly inhibits many extracellular matrix genes, a function that is conserved in mammals (Whetstine et al., 2005). These C. elegans studies have thus begun to identify gene regulatory networks that are controlled through particular modifications of histones or other transcription factors, and may have been broadly conserved during evolution.
Much remains to be learned about the functions of CBP-1 and HDA-1, but it is clear that they are involved in additional biological and regulatory pathways of major importance. While the above processes appear to involve maternally-provided CBP-1, impairment of zygotically-derived CBP-1 results in embryonic arrest at the beginning of morphogenesis, indicating that CBP-1 is needed for additional developmental processes (Victor et al., 2002). HDA-1 has been implicated in additional functions that include vulval development and suppression of inappropriate germline gene expression (Dufourcq et al., 2002; Unhavaithaya et al., 2002). These HDA-1 functions involve a genetic pathway (SynMuvB) that negatively regulates specification of the vulval cell fate, and includes other conserved chromatin modification factors along with LIN-35, the C. elegans ortholog of the retinoblastoma tumor suppressor protein (Fay and Han, 2000; Lu and Horvitz, 1998). Genetic and RNAi screening has identified a surprisingly intricate network of additional chromatin modifiers and other regulatory factors that function either to cooperate with or oppose the action of SynMuv pathway components in vulval fate specification (Cui et al., 2006b; Poulin et al., 2005). Further illustrating the complexity of these functional interactions, CBP-1 appears to antagonize vulval fate specification by a mechanism that is independent of the SynMuv pathway (Eastburn and Han, 2005). Recent work indicates that the SynMuv pathway may function by repressing hypodermal expression of the Epidermal Growth Factor-related protein LIN-3, which drives the RAS/MAPK signal that induces vulval cell fate (Cui et al., 2006a). This last finding may eventually provide an entry point for mechanistic analyses of how particular chromatin modifiers interact functionally with other transcription factors to regulate expression of this key developmental gene. These studies of how chromatin modification machines regulate vulval cell fate illustrate the power of C. elegans genetics for discovering functional relationships that can be interpreted in the light of known biochemical functions, and can lead to new insights into regulation of specific biological processes in vivo.
Development of an organism depends upon finely-tuned and accurate control of mRNA transcription. Recent studies have identified various biological processes in C. elegans that involve direct regulation or specific functions of general transcription machinery components such as Mediator, or transcription co-factors that function in multiple discrete contexts, such as chromatin modification proteins. The list of these processes will continue to grow, as future studies identify additional examples of direct communications between regulatory and general transcription factors, and reveal how promoter groups and gene networks are regulated coordinately through these interactions. Further analyses of these regulatory mechanisms in C. elegans should continue to provide broadly applicable information about their conserved functions in vivo, and new insights into the specific biological processes in which they are involved.
Table 1. C. elegans genes that encode major GTFs. These factors are listed according to their names as defined in biochemical or yeast genetic studies, except for TAFIIs and Mediator subunits, for which unified nomenclatures have been agreed upon (Tora, 2004; Bourbon et al., 2004). C. elegans TAFIIs were previously named according to an earlier nomenclature (Walker et al., 2001). Each of these C. elegans genes was identified as the indicated GTF through homology searches performed either in published studies (Worm Base) or by the authors. Blanks in the C. elegans column indicate factors for which no ortholog has yet been identified.
| Complex | Name | Former name | C. elegans gene | ORF |
|---|---|---|---|---|
| RNA Pol II | Rpb1 | ama-1 | F36A4.7 | |
| Rpb2 | rpb-2 | C26E6.4 | ||
| Rpb3 | C36B1.3 | |||
| Rpb4 | F43E2.2 | |||
| Rpb5 | H27M09.2 | |||
| Rpb6 | C06A1.5 | |||
| Rpb7 | Y54E10BR.6 | |||
| Rpb8 | F26F4.11 | |||
| Rpb9 | Y97E10AR.5 | |||
| Rpb10 | Y37E3.3 | |||
| Rpb11 | W01G7.3 | |||
| Rpb12 | F23B2.13 | |||
| TFIIA | alpha/beta subunit | pqn-51 | K11D12.2 | |
| TFIIA | gamma subunit | Y111B2A.13 | ||
| TFIIB | ttb-1 | W03F9.5 | ||
| TFIID | TBP | tbp-1 | T20B12.2 | |
| TAF1 | taf-1 | W04A8.7 | ||
| TAF 2 | taf-2 | Y37E11B.4 | ||
| TAF 3 | taf-3 | C11G6.1 | ||
| TAF 4 | taf-5 | taf-4 | R119.6 | |
| TAF 5 | taf-4 | taf-5 | F30F8.8 | |
| TAF 6 | taf-3.1 | taf-6.1 | W09B6.2 | |
| TAF 6L | taf-3.2 | taf-6.2 | Y37E11AL.8 | |
| TAF 7 | taf-8.1 | taf-7.1 | F54F7.1 | |
| TAF 7L | taf-8.2 | taf-7.2 | Y111B2A.16 | |
| TAF 8 | taf-8 | ZK1320.12 | ||
| TAF 9 | taf-10 | taf-9 | T12D8.7 | |
| TAF 10 | taf-11 | taf-10 | K03B4.3 | |
| TAF 11 | taf-7.1 | taf-11.1 | F48D6.1 | |
| TAF 11L | taf-7.2 | taf-11.2 | K10D3.3 | |
| TAF12 | taf-9 | taf-12 | Y56A3A.4 | |
| TAF13 | taf-6 | taf-13 | C14A4.10 | |
| TAF14 | ||||
| TAF15 | ||||
| TFIIE | alpha subunit | F54D5.11 | ||
| beta subunit | ||||
| TFIIF | large subunit | C01F1.1 | ||
| TFIIF | small subunit | Y39B6A.36 | ||
| TFIIH | XPB | Y66D12A.15 | ||
| XPD | Y50D7A.2 | |||
| p62 | R02D3.3 | |||
| p52 | ||||
| p44 | T16h12.4 | |||
| p34 | ZK1128.4 | |||
| CDK7 | cdk-7 | Y39G10AL.3 | ||
| CyclinH | cyh-1 | Y49F6B.1 | ||
| MAT1 | mnat-1 | F53g2.7 | ||
| Mediator | MED1 | sop-3 | mdt-1.1 | Y71F9B.10 |
| mdt-1.2 | T23C6.1 | |||
| MED2 | ||||
| MED3 | ||||
| MED4 | mdt-4 | ZK546.13 | ||
| MED5 | ||||
| MED6 | let-425/med-6 | mdt-6 | Y57E12AL.5 | |
| MED7 | let-49/med-7 | mdt-7 | Y54E5B.3 | |
| MED8 | mdt-8 | Y62F5A.1b | ||
| MED9 | ||||
| MED10 | mdt-10 | T09A5.6 | ||
| MED11 | mdt-11 | R144.9 | ||
| MED12 | dpy-22/sop-1 | mdt-12 | F47A4.2 | |
| MED13 | let-19 | mdt-13 | K08F8.6 | |
| MED14 | rgr-1 | mdt-14 | C38C10.5 | |
| MED15 | mdt-15 | R12B2.5b | ||
| MED16 | ||||
| MED17 | mdt-17 | Y113G7B.18 | ||
| MED18 | mdt-18 | C55B7.9 | ||
| MED19 | mdt-19 | Y71H2B.6 | ||
| MED20 | mdt-20 | Y104H12D.1 | ||
| MED21 | mdt-21 | C24H11.9 | ||
| MED22 | mdt-22 | ZK970.3 | ||
| MED23 | sur-2 | mdt-23 | F39B2.4 | |
| MED24 | ||||
| MED25 | ||||
| MED26 | ||||
| MED27 | mdt-27 | T18H9.6 | ||
| MED28 | mdt-28 | W01A8.1 | ||
| MED29 | mdt-29 | K08E3.8 | ||
| MED30 | ||||
| MED31 | mdt-31 | F32H2.2 | ||
| CDK8 | cdk-8 | F39H11.3 | ||
| CycC | cic-1 | H14E04.5 | ||
| PTEFb | CycT | cit-1.1 | F44B9.4 | |
| cit-1.2 | F44B9.3 | |||
| CDK9 | cdk-9 | H25P06.2 | ||
| Other | TLF | tlf-1 | F39H11.2 |
This work was supported by a grant from the National Institute of General Medical Sciences (GM63826).
Batchelder, C., Dunn, M.A., Choy, B., Suh, Y., Cassie, C., Shim, E.Y., Shin, T.H., Mello, C., Seydoux, G., and Blackwell, T.K. (1999). Transcriptional repression by the C. elegans germline protein PIE-1. Genes Dev. 13, 202–212. Abstract
Baugh, L.R., Hill, A.A., Slonim, D.K., Brown, E.L., and Hunter, C.P. (2003). Composition and dynamics of the Caenorhabditis elegans early embryonic transcriptome. Development 130, 889–900. Abstract Article
Bentley, D.L. (2005). Rules of engagement: co-transcriptional recruitment of pre-mRNA processing factors. Curr. Opin. Cell Biol. 17, 251–256. Abstract Article
Bird, D.M., and Riddle, D.L. (1989). Molecular cloning and sequencing of ama-1, the gene encoding the largest subunit of Caenorhabditis elegans RNA polymerase II. Mol. Cell. Biol. 9, 4119–4130. Abstract
Bjorklund, S., and Gustafsson, C.M. (2005). The yeast Mediator complex and its regulation. Trends Biochem. Sci. 30, 240–244. Abstract Article
Bourbon, H.M., Aguilera, A., Ansari, A.Z., Asturias, F.J., Berk, A.J., Bjorklund, S., Blackwell, T.K., Borggrefe, T., Carey, M., Carlson, M., et al. (2004). A unified nomenclature for protein subunits of mediator complexes linking transcriptional regulators to RNA polymerase II. Mol. Cell 14, 553–557. Abstract Article
Boyer, T.G., Martin, M.E., Lees, E., Ricciardi, R.P., and Berk, A.J. (1999). Mammalian Srb/Mediator complex is targeted by adenovirus E1A protein. Nature 399, 276–279. Abstract Article
Buratowski, S. (2005). Connections between mRNA 3′ end processing and transcription termination. Curr. Opin. Cell Biol. 17, 257–261. Abstract Article
Calvo, D., Victor, M., Gay, F., Sui, G., Luke, M.P., Dufourcq, P., Wen, G., Maduro, M., Rothman, J., and Shi, Y. (2001). A POP-1 repressor complex restricts inappropriate cell type-specific gene transcription during Caenorhabditis elegans embryogenesis. EMBO J. 20, 7197–7208. Abstract Article
Chan, H.M., and La Thangue, N.B. (2001). p300/CBP proteins: HATs for transcriptional bridges and scaffolds. J. Cell. Sci. 114, 2363–2373. Abstract
Chong, J.A., Moran, M.M., Teichmann, M., Kaczmarek, J.S., Roeder, R., and Clapham, D.E. (2005). TATA-binding protein (TBP)-like factor (TLF) is a functional regulator of transcription: reciprocal regulation of the neurofibromatosis type 1 and c-fos genes by TLF/TRF2 and TBP. Mol. Cell. Biol. 25, 2632–2643. Abstract Article
Chu, D.S., Dawes, H.E., Lieb, J.D., Chan, R.C., Kuo, A.F., and Meyer, B.J. (2002). A molecular link between gene-specific and chromosome-wide transcriptional repression. Genes Dev. 16, 796–805. Abstract Article
Cui, M., Chen, J., Myers, T.R., Hwang, B.J., Sternberg, P.W., Greenwald, I., and Han, M. (2006a). SynMuv genes redundantly inhibit lin-3/EGF expression to prevent inappropriate vulval induction in C. elegans. Dev. Cell 10, 667–672. Abstract Article
Cui, M., Kim, E.B., and Han, M. (2006b). Diverse chromatin remodeling genes antagonize the Rb-involved SynMuv pathways in C. elegans. PLoS Genet. 2, e74. Abstract Article
Dantonel, J.C., Quintin, S., Lakatos, L., Labouesse, M., and Tora, L. (2000). TBP-like factor is required for embryonic RNA polymerase II transcription in C. elegans. Mol. Cell 6, 715–722. Abstract Article
Deplancke, B., Mukhopadhyay, A., Ao, W., Elewa, A.M., Grove, C.A., Martinez, N.J., Sequerra, R., Doucette-Stamm, L., Reece-Hoyes, J.S., Hope, I.A., et al. (2006). A gene-centered C. elegans protein-DNA interaction network. Cell 125, 1193–1205. Abstract Article
Dufourcq, P., Victor, M., Gay, F., Calvo, D., Hodgkin, J., and Shi, Y. (2002). Functional requirement for histone deacetylase 1 in Caenorhabditis elegans gonadogenesis. Mol. Cell. Biol. 22, 3024–3034. Abstract Article
Dupuy, D., Li, Q.R., Deplancke, B., Boxem, M., Hao, T., Lamesch, P., Sequerra, R., Bosak, S., Doucette-Stamm, L., Hope, I.A., et al. (2004). A first version of the Caenorhabditis elegans Promoterome. Genome Res. 14, 2169–2175. Abstract Article
Eastburn, D.J., and Han, M. (2005). A gain-of-function allele of cbp-1, the Caenorhabditis elegans ortholog of the mammalian CBP/p300 gene, causes an increase in histone acetyltransferase activity and antagonism of activated Ras. Mol. Cell. Biol. 25, 9427–9434. Abstract Article
Edgar, L.G., Wolf, N., and Wood, W.B. (1994). Early transcription in Caenorhabditis elegans embryos. Development 120, 443–451. Abstract
Fay, D.S., and Han, M. (2000). The synthetic multivulval genes of C. elegans: functional redundancy, Ras-antagonism, and cell fate determination. Genesis 26, 279–284. Abstract
Gibert, M.A., Starck, J., and Beguet, B. (1984). Role of the gonad cytoplasmic core during oogenesis of the nematode Caenorhabditis elegans. Biol. Cell. 50, 77–85. Abstract
Green, M.R. (2005). Eukaryotic transcription activation: right on target. Mol. Cell 18, 399–402. Abstract Article
Hochheimer, A., Zhou, S., Zheng, S., Holmes, M.C., and Tjian, R. (2002). TRF2 associates with DREF and directs promoter-selective gene expression in Drosophila. Nature 420, 439–445. Abstract Article
Huisinga, K.L., and Pugh, B.F. (2004). A genome-wide housekeeping role for TFIID and a highly regulated stress-related role for SAGA in Saccharomyces cerevisiae. Mol. Cell 13, 573–585. Abstract Article
Kaltenbach, L., Horner, M.A., Rothman, J.H., and Mango, S.E. (2000). The TBP-like factor CeTLF is required to activate RNA polymerase II transcription during C. elegans embryogenesis. Mol. Cell 6, 705–713. Abstract Article
Kelly, W.G., Schaner, C.E., Dernburg, A.F., Lee, M.H., Kim, S.K., Villeneuve, A.M., and Reinke, V. (2002). X-chromosome silencing in the germline of C. elegans. Development 129, 479–492. Abstract
Kornberg, R.D. (2005). Mediator and the mechanism of transcriptional activation. Trends Biochem. Sci. 30, 235–239. Abstract Article
Kwon, J.Y., and Lee, J. (2001). Biological significance of a universally conserved transcription mediator in metazoan developmental signaling pathways. Development 128, 3095–3104. Abstract
Kwon, J.Y., Park, J.M., Gim, B.S., Han, S.J., Lee, J., and Kim, Y.J. (1999). Caenorhabditis elegans mediator complexes are required for developmental-specific transcriptional activation. Proc. Natl. Acad. Sci. U.S.A. 96, 14990–14995. Abstract Article
Lemon, B., and Tjian, R. (2000). Orchestrated response: a symphony of transcription factors for gene control. Genes Dev. 14, 2551–2569. Abstract Article
Lichtsteiner, S., and Tjian, R. (1993). Cloning and properties of the Caenorhabditis elegans TATA-box-binding protein. Proc. Natl. Acad. Sci. U.S.A. 90, 9673–9677. Abstract Article
Lichtsteiner, S., and Tjian, R. (1995). Synergistic activation of transcription by UNC-86 and MEC-3 in Caenorhabditis elegans embryo extracts. EMBO J. 14, 3937–3945. Abstract
Lu, X., and Horvitz, H.R. (1998). lin-35 and lin-53, two genes that antagonize a C. elegans Ras pathway, encode proteins similar to Rb and its binding protein RbAp48. Cell 95, 981–991. Abstract Article
Malik, S., and Roeder, R.G. (2005). Dynamic regulation of pol II transcription by the mammalian Mediator complex. Trends Biochem. Sci. 30, 256–263. Abstract Article
Martianov, I., Brancorsini, S., Gansmuller, A., Parvinen, M., Davidson, I., and Sassone-Corsi, P. (2002). Distinct functions of TBP and TLF/TRF2 during spermatogenesis: requirement of TLF for heterochromatic chromocenter formation in haploid round spermatids. Development 129, 945–955. Abstract
Martianov, I., Fimia, G.M., Dierich, A., Parvinen, M., Sassone-Corsi, P., and Davidson, I. (2001). Late arrest of spermiogenesis and germ cell apoptosis in mice lacking the TBP-like TLF/TRF2 gene. Mol. Cell 7, 509–515. Abstract Article
Meinhart, A., Kamenski, T., Hoeppner, S., Baumli, S., and Cramer, P. (2005). A structural perspective of CTD function. Genes Dev. 19, 1401–1415. Abstract Article
Mello, C.C., Draper, B.W., Krause, M., Weintraub, H., and Priess, J.R. (1992). The pie-1 and mex-1 genes and maternal control of blastomere identity in early C. elegans embryos. Cell 70, 163–176. Abstract Article
Moghal, N., and Sternberg, P.W. (2003). A component of the transcriptional mediator complex inhibits RAS-dependent vulval fate specification in C. elegans. Development 130, 57–69. Abstract Article
Muller, F., Lakatos, L., Dantonel, J., Strahle, U., and Tora, L. (2001). TBP is not universally required for zygotic RNA polymerase II transcription in zebrafish. Curr. Biol. 11, 282–287. Abstract Article
Muller, F., and Tora, L. (2004). The multicoloured world of promoter recognition complexes. EMBO J. 23, 2–8. Abstract Article
Oh, S.W., Mukhopadhyay, A., Dixit, B.L., Raha, T., Green, M.R., and Tissenbaum, H.A. (2006). Identification of direct DAF-16 targets controlling longevity, metabolism and diapause by chromatin immunoprecipitation. Nat. Genet. 38, 251–257. Abstract Article
Orphanides, G., and Reinberg, D. (2002). A unified theory of gene expression. Cell 108, 439–451. Abstract Article
Poulin, G., Dong, Y., Fraser, A.G., Hopper, N.A., and Ahringer, J. (2005). Chromatin regulation and sumoylation in the inhibition of Ras-induced vulval development in Caenorhabditis elegans. EMBO J. 24, 2613–2623. Abstract Article
Powell-Coffman, J.A., Knight, J., and Wood, W.B. (1996). Onset of C. elegans gastrulation is blocked by inhibition of embryonic transcription with an RNA polymerase antisense RNA. Dev. Biol. 178, 472–483. Abstract Article
Price, D.H. (2000). P-TEFb, a cyclin-dependent kinase controlling elongation by RNA polymerase II. Mol. Cell. Biol. 20, 2629–2634. Abstract Article
Rogalski, T.M., Bullerjahn, A.M., and Riddle, D.L. (1988). Lethal and amanitin-resistance mutations in the Caenorhabditis elegans ama-1 and ama-2 genes. Genetics 120, 409–422. Abstract
Sanford, T., Golomb, M., and Riddle, D.L. (1983). RNA polymerase II from wild type and alpha-amanitin-resistant strains of Caenorhabditis elegans. J. Biol. Chem. 258, 12804–12809. Abstract
Sanford, T., Prenger, J.P., and Golomb, M. (1985). Purification and immunological analysis of RNA polymerase II from Caenorhabditis elegans. J. Biol. Chem. 260, 8064–8069. Abstract
Schaner, C.E., Deshpande, G., Schedl, P.D., and Kelly, W.G. (2003). A conserved chromatin architecture marks and maintains the restricted germ cell lineage in worms and flies. Dev. Cell 5, 747–757. Abstract Article
Schisa, J.A., Pitt, J.N., and Priess, J.R. (2001). Analysis of RNA associated with P granules in germ cells of C. elegans adults. Development 128, 1287–1298. Abstract
Seydoux, G., and Dunn, M.A. (1997). Transcriptionally repressed germ cells lack a subpopulation of phosphorylated RNA polymerase II in early embryos of Caenorhabditis elegans and Drosophila melanogaster. Development 124, 2191–2201. Abstract
Seydoux, G., and Fire, A. (1994). Soma-germline asymmetry in the distributions of embryonic RNAs in Caenorhabditis elegans. Development 120, 2823–2834. Abstract
Seydoux, G., Mello, C.C., Pettitt, J., Wood, W.B., Priess, J.R., and Fire, A. (1996). Repression of gene expression in the embryonic germ lineage of C. elegans. Nature 382, 713–716. Abstract Article
Shi, Y., and Mello, C. (1998). A CBP/p300 homolog specifies multiple differentiation pathways in Caenorhabditis elegans. Genes Dev. 12, 943–955. Abstract
Shim, E.Y., Walker, A.K., and Blackwell, T.K. (2002a). Broad requirement for the Mediator subunit RGR-1 for transcription in the C. elegans embryo. J. Biol. Chem. 277, 30413–30416. Abstract Article
Shim, E.Y., Walker, A.K., Shi, Y., and Blackwell, T.K. (2002b). CDK-9/Cyclin T (P-TEFb) is required in two post-initiation pathways for transcription in the C. elegans embryo. Genes Dev. 16, 2135–2146. Abstract Article
Singh, N., and Han, M. (1995). sur-2, a novel gene, functions late in the let-60 ras-mediated signaling pathway during Caenorhabditis elegans vulval induction. Genes Dev. 9, 2251–2265. Abstract
Smale, S.T., and Kadonaga, J.T. (2003). The RNA polymerase II core promoter. Annu. Rev. Biochem. 72, 449–479. Abstract Article
Stevens, J.L., Cantin, G.T., Wang, G., Shevchenko, A., and Berk, A.J. (2002). Transcription control by E1A and MAP kinase pathway via Sur2 mediator subunit. Science 296, 755–758. Abstract Article
Takagi, T., Walker, A.K., Sawa, C., Diehn, F., Takase, Y., Blackwell, T.K., and Buratowski, S. (2003). The Caenorhabditis elegans mRNA 5′-Capping Enzyme. IN VITRO AND IN VIVO CHARACTERIZATION. J. Biol. Chem. 278, 14174–14184. Abstract Article
Taubert, S., Van Gilst, M.R., Hansen, M., and Yamamoto, K.R. (2006). A Mediator subunit, MDT-15, integrates regulation of fatty acid metabolism by NHR-49-dependent and -independent pathways in C. elegans. Genes Dev. 20, 1137–1149. Abstract Article
Tenenhaus, C., Subramaniam, K., Dunn, M.A., and Seydoux, G. (2001). PIE-1 is a bifunctional protein that regulates maternal and zygotic gene expression in the embryonic germ line of Caenorhabditis elegans. Genes Dev. 15, 1031–1040. Abstract Article
Tora, L. (2002). A unified nomenclature for TATA box binding protein (TBP)-associated factors (TAFs) involved in RNA polymerase II transcription. Genes Dev. 16, 673–675. Abstract Article
Unhavaithaya, Y., Shin, T.H., Miliaras, N., Lee, J., Oyama, T., and Mello, C.C. (2002). MEP-1 and a homolog of the NURD complex component Mi-2 act together to maintain germline-soma distinctions in C. elegans. Cell 111, 991–1002. Abstract Article
Veenstra, G.J., Weeks, D.L., and Wolffe, A.P. (2000). Distinct roles for TBP and TBP-like factor in early embryonic gene transcription in Xenopus. Science 290, 2312–2315. Abstract Article
Victor, M., Bei, Y., Gay, F., Calvo, D., Mello, C., and Shi, Y. (2002). HAT activity is essential for CBP-1-dependent transcription and differentiation in Caenorhabditis elegans. EMBO Rep. 3, 50–55. Abstract Article
Walker, A.K., and Blackwell, T.K. (2003). A Broad but Restricted Requirement for TAF-5 (Human TAFII100) for Embryonic Transcription in Caenorhabditis elegans. J. Biol. Chem. 278, 6181–6186. Abstract Article
Walker, A.K., Rothman, J.H., Shi, Y., and Blackwell, T.K. (2001). Distinct requirements for C.elegans TAF(II)s in early embryonic transcription. EMBO J. 20, 5269–5279. Abstract Article
Walker, A.K., Shi, Y., and Blackwell, T.K. (2004). An extensive requirement for TFIID-specific TAF-1 in C. elegans embryonic transcription. J. Biol. Chem. 279, 29270–29277. Abstract Article
Wallenfang, M.R., and Seydoux, G. (2002). cdk-7 is required for mRNA transcription and cell cycle progression in C. elegans embryos. Proc. Natl. Acad. Sci. U.S.A. 99, 5527–5532. Abstract Article
Wang, G., Balamotis, M.A., Stevens, J.L., Yamaguchi, Y., Handa, H., and Berk, A.J. (2005). Mediator requirement for both recruitment and postrecruitment steps in transcription initiation. Mol. Cell 17, 683–694. Abstract Article
Wang, J.C., Walker, A., Blackwell, T.K., and Yamamoto, K.R. (2004). The Caenorhabditis elegans ortholog of TRAP240, CeTRAP240/let-19, selectively modulates gene expression and is essential for embryogenesis. J. Biol. Chem. 279, 29270–29277. Abstract Article
Whetstine, J.R., Ceron, J., Ladd, B., Dufourcq, P., Reinke, V., and Shi, Y. (2005). Regulation of tissue-specific and extracellular matrix-related genes by a class I histone deacetylase. Mol. Cell 18, 483–490. Abstract Article
Yang, F., Vought, B.W., Satterlee, J.S., Walker, A.K., Jim Sun, Z.Y., Watts, J.L., Debeaumont, R., Mako Saito, R., Hyberts, S.G., Yang, S., et al. (2006). An ARC/Mediator subunit required for SREBP control of cholesterol and lipid homeostasis. Nature 442, 700–704. Abstract Article
Yoda, A., Kouike, H., Okano, H., and Sawa, H. (2005). Components of the transcriptional Mediator complex are required for asymmetric cell division in C. elegans. Development 132, 1885–1893. Abstract Article
Zhang, F., Barboric, M., Blackwell, T.K., and Peterlin, B.M. (2003). A model of repression: CTD analogs and PIE-1 inhibit transcriptional elongation by P-TEFb. Genes Dev. 17, 748–758. Abstract Article
Zhang, H., and Emmons, S.W. (2000). A C. elegans mediator protein confers regulatory selectivity on lineage-specific expression of a transcription factor gene. Genes Dev. 14, 2161–2172. Abstract Article
Zhang, H., and Emmons, S.W. (2001). The novel C. elegans gene sop-3 modulates Wnt signaling to regulate Hox gene expression. Development 128, 767–777. Abstract
*Edited by Thomas Blumenthal. Last revised July 14, 2006. Published September 5, 2006. This chapter should be cited as: Blackwell, T. K. and Walker, A. K. Transcription mechanisms (September 5, 2006), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.121.1, http://www.wormbook.org.
Copyright: © 2006 T. Keith Blackwell and Amy K. Walker. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
§To whom correspondence should be addressed. Phone: 617-919-2769, E-mail: keith.blackwell@joslin.harvard.edu
†Present Address: Massachusetts General Hospital Cancer Center, 149-7202 13th Street, Charlestown, MA 02129
 All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.
All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.