Embryo series courtesy of Einhard Schierenberg
Embryo series courtesy of Einhard SchierenbergTable of Contents
Abstract
Receptor Tyrosine Kinase (RTK)/Ras GTPase/MAP kinase (MAPK) signaling pathways are used repeatedly during metazoan development to control many different biological processes. In the nematode Caenorhabditis elegans, two different RTKs (LET-23/EGFR and EGL-15/FGFR) are known to stimulate LET-60/Ras and a MAPK cascade consisting of the kinases LIN-45/Raf, MEK-2/MEK and MPK-1/ERK. This Ras/MAPK cascade is required for multiple developmental events, including induction of vulval, uterine, spicule, P12 and excretory duct cell fates, control of sex myoblast migration and axon guidance, and promotion of germline meiosis. Studies in C. elegans have provided much insight into the basic framework of this RTK/Ras/MAPK signaling pathway, its regulation, how it elicits cell-type specific responses, and how it interacts with other signaling pathways such as the Wnt and Notch pathways.
Receptor Tyrosine Kinase (RTK)/Ras GTPase/MAP kinase (MAPK) signaling pathways are used repeatedly during metazoan development to control many different biological processes (Schlessinger, 2000). Mutations affecting RTK/Ras/MAPK signaling cause many human syndromes and diseases, including cancer (Malumbres and Barbacid, 2002). Studies in C. elegans have provided much insight into the basic framework of this canonical type of Ras pathway, its regulation, how it elicits cell-type specific responses, and how it interacts with other signaling pathways (for recent reviews see Moghal and Sternberg, 2003; Sundaram, 2004; Tan and Kim, 1999; Wang and Sternberg, 2001).
C. elegans Ras is called LET-60 (Han and Sternberg, 1990). LET-60 Ras acts downstream of at least two different RTKs, LET-23 (related to the Epidermal Growth Factor Receptor or EGFR; Aroian et al., 1990) and EGL-15 (related to the Fibroblast Growth Factor Receptor or FGFR; DeVore et al., 1995). The only known role of LET-60 Ras is to stimulate a MAPK cascade consisting of the kinases LIN-45 (Raf; Han et al., 1993), MEK-2 (MEK; Church et al., 1995; Kornfeld et al., 1995; Wu et al., 1995) and MPK-1 (ERK/MAPK; Lackner et al., 1994; Wu and Han, 1994). LET-60 Ras signaling is required for multiple developmental events, the best studied of which is vulval induction (see Vulval development). Genetic screens based on various let-60 mutant phenotypes have identified many generally-acting “core” components of the Ras pathway as well as numerous regulators or targets of the pathway (Figure 1; Table 1). Indeed, many important Ras pathway genes were first identified in the worm.
|
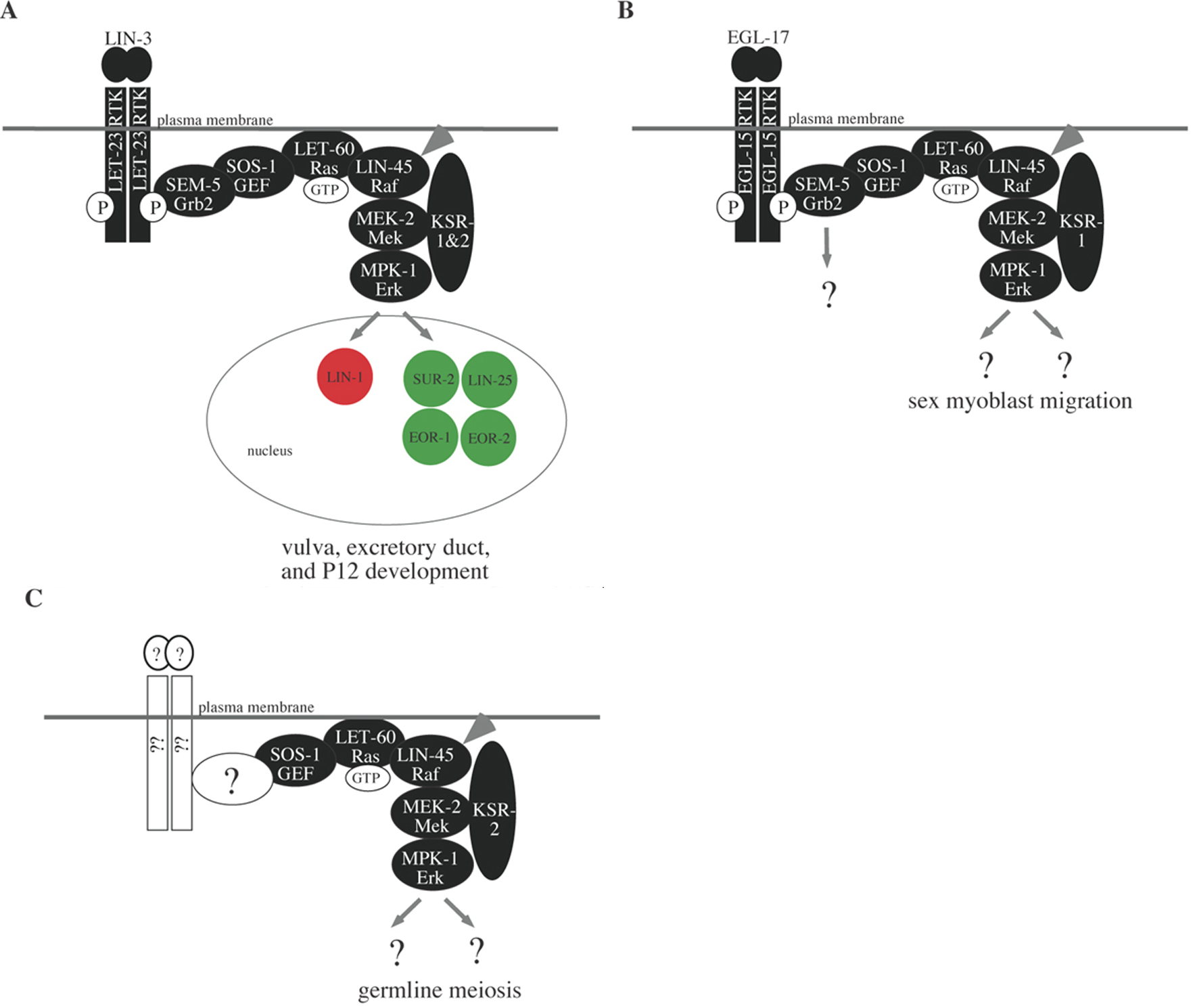
Figure 1. Variations of the RTK/Ras/MAPK signaling pathway controlling different developmental processes in C. elegans. The RTKs LET-23 (EGFR) and EGL-15 (FGFR) are activated by different ligands and control different sets of developmental processes. A) LIN-3/LET-23-dependent processes include vulval, excretory duct and P12 development; B) EGL-17/EGL-15-dependent processes include sex myoblast migration; C) Neither LET-23 nor EGL-15 control pachytene progression during germline meiosis. Both RTKs signal through the adaptor SEM-5 (Grb2) to activate the same core Ras/MAPK pathway. EGL-15 and SEM-5 may also activate additional pathways during sex myoblast migration. The scaffold proteins KSR-1 and KSR-2 assist in LIN-45 (Raf) and/or MEK-2 activation; different processes have different requirements for these two KSR proteins. Shown in A) are five nuclear proteins (LIN-1, SUR-2, LIN-25, EOR-1, EOR-2) that are jointly important for vulval, excretory duct and P12 cell fates. Downstream targets of MPK-1 during sex myoblast migration and germline meiosis are not yet known. See text and Table 1 for details and references.
Table 1. Core components, regulators and targets of the C. elegans RTK/Ras/MAPK signaling pathway. Core components are shown in black, positive regulators or targets are shown in green, and negative regulators or targets are shown in red.
| Gene product | Mammalian relative(s) | Molecular function | Ras-related phenotypes | Reference(s) | |
|---|---|---|---|---|---|
| ARK-1 | Ack-Related Kinase | Ack | Tyrosine kinase | ~WT. Muv in combination with sli-1, others. | Hopper et al., 2000 |
| CDF-1 | Cation Diffusion Facilitator | ZnT1 | Zinc transporter. | ~WT. Suppresses let-60 ras(gf) Muv. | Bruinsma et al., 2002; Jakubowski and Kornfeld, 1999 |
| CNK-1 | Connector/eNhancer of Ksr | CNK1,2,3 | Putative Raf-binding adaptor/scaffold. | ~WT. Suppresses let-60 ras(gf) Muv. Enhances Vul and rod-like lethal phenotypes of lin-45, others. | Rocheleau et al., 2005 |
| DAB-1 | DisABled homolog | DAB2 | Adaptor. Required for EGL-17 secretion. | Abnormal sex myoblast positions. | Kamikura and Cooper, 2003 |
| DPY-22/ SOP-1 | DumPY/Suppressor Of Pal-1 | TRAP230 | Mediator subunit. | ~WT. Enhances let-23(gf) Muv. | Moghal and Sternberg, 2003 |
| DPY-23 | DumPY | AP-50 | Clathrin adaptor subunit | ~WT. Muv in combination with gap-1. | Yoo et al., 2004 |
| EGL-5 | EGg-Laying defects | Hox9-13 | Hox transcription factor. Upregulated by Ras signaling. | P12→P11 fate transformations. | Chisholm, 1991; Jiang and Sternberg, 1998 |
| EGL-18 | EGg-Laying defects | GATA4,5,6 | GATA transcription factor | Partial Vul. Strong Vul in combination with elt-6. | Koh et al., 2002; Koh et al., 2004 |
| EGL-15 | EGg Laying defects | FGFR | Receptor tyrosine kinase | Larval lethal. Abnormal sex myoblast positions. Axon guidance defects. | Bulow et al., 2004; DeVore et al., 1995; Goodman et al., 2003; Stern and Horvitz 1991 |
| EGL-17 | EGg-Laying defects | FGF | Ligand for EGL-15 RTK | Abnormal sex myoblast positions. | Burdine et al., 1997; Stern and Horvitz 1991 |
| EGL-19 | Egg-Laying defects | Cav1.2 | Alpha subunit of L-type voltage-gated calcium channel | Suppresses egl-30(gf) effects on vulva | Moghal and Sternberg, 2003 |
| EGL-30 | EGg-Laying defects | Gαq/Gα11 | Galphaq subunit of heterotrimeric G protein | gf allele suppresses let-23 and let-60(dn) Vul | Moghal and Sternberg, 2003 |
| ELT-6 | Erythroid-Like Transcription factor family | GATA4,5,6 | GATA transcription factor | ~WT. Vul in combination with egl-18. | Koh et al., 2002; Koh et al., 2004 |
| EOR-1 | Egl-1 suppressor/diO uptake defective/ Rafenhancer | PLZF | BTB/Zinc finger protein, probable transcriptional regulator | Partial rod-like lethal and P12→P11 fate changes. Strong rod-like lethal in combination with sur-2, lin-25 or lin-1. | Howard and Sundaram, 2002; Rocheleau et al., 2002 |
| EOR-2 | Egl-1 suppressor/diO uptake defective/ Rafenhancer | NP_079001.2 | Novel nuclear protein, functions with EOR-1. | Partial rod-like lethal and P12→P11 fate changes. Strong rod-like lethal in combination with sur-2, lin-25 or lin-1. | Howard and Sundaram, 2002; Rocheleau et al., 2002 |
| GAP-1 | GTPase Activating Protein | GAP-1 | Ras GAP | ~WT. Suppresses let-23 Vul. Muv in combination with lip-1, others. | Hajnal et al., 1997 |
| GAP-2 | GTPase Activating Protein | p120GAP | Ras GAP | ~WT. Suppresses let-23 rod-like lethality. | Hayashizaki et al., 1998 |
| GPA-5 | G-Protein, Alpha subunit | GNAZ | Ga | Increased chemotaxis. Suppresses let-60(dn) Vul. | Battu et al, 2003 |
| KSR-1 | Kinase Suppressor of Ras | KSR1, 2 | Raf-related MEK-binding protein, scaffold for Raf/MEK/ERK | Abnormal sex myoblast positions. Suppresses let-60 ras(gf) Muv. Rod-like lethal and Vul in combination with ksr-2, others. | Kornfeld et al., 1995b; Sundaram and Han, 1995 |
| KSR-2 | Kinase Suppressor of Ras | KSR1, 2 | Raf-related MEK-binding protein, scaffold for Raf/MEK/ERK | Sterile. Rod-like lethal and Vul in combination with ksr-1. | Ohmachi et al., 2002 |
| LET-23 | LEThal | EGFR | Receptor tyrosine kinase | Rod-like larval lethal, Vul, etc. | Aroian et al., 1990 |
| LET-60 | LEThal | K-Ras | Small GTPase | Rod-like larval lethal, Vul, sterile, etc. | Beitel et al., 1990; Han et al., 1990; Han and Sternberg, 1990 |
| LET-92 | LEThal | PPP2CB | Catalytic subunit of Protein Phosphatase 2A | Dominantly enhances let-60(dn) Vul | Kao et al., 2004 |
| LET-756 | LEThal | FGF9 | Ligand for EGL-15 RTK | Larval lethal. Axon guidance defects. | Bulow et al., 2004; Popovici et al., 2004; Roubin et al., 1999 |
| LIN-1 | Abnormal cell LINeage | Elk1 | Ets domain transcription factor. Target of MPK-1. | Muv | Beitel et al., 1995; Jacobs et al., 1998 |
| LIN-2 | Abnormal cell LINeage | CASK | Membrane-associated Guanylate kinase, required for basal localization of LET-23 | Vul | Hoskins et al., 1996; Kaech et al., 1998 |
| LIN-3 | Abnormal cell LINeage | EGF | Ligand for LET-23 RTK | Rod-like larval lethal, Vul, etc. | Hill and Sternberg, 1992; Dutt et al., 2004 |
| LIN-7 | Abnormal cell LINeage | Lin-7 | PDZ and PTB-domain protein, required for basal localization of LET-23 | Vul | Kaech et al., 1998; Simske et al., 1996 |
| LIN-10 | Abnormal cell LINeage | Mint1, 2, 3 | PDZ protein, required for basal localization of LET-23 | Vul | Kaech et al., 1998; Whitfield et al., 1999 |
| LIN-25 | Abnormal cell LINeage | - | Novel nuclear protein, functions with SUR-2 | Vul | Nilsson et al., 1998; Nilsson et al., 2000; Tuck and Greenwald, 1995 |
| LIN-31 | Abnormal cell LINeage | FoxB2 | Winged helix transcription factor. Target of MPK-1. | Mixed Vul and Muv | Miller et al., 1993; Tan et al., 1998 |
| LIN-39 | Abnormal cell LINeage | HoxB5 | Hox transcription factor. Upregulated by Ras signaling. | Vul | Clark et al., 1993; Maloof and Kenyon, 1998 |
| LIN-45 | Abnormal cell LINeage | B-Raf | Serine/threonine kinase. Binds Ras-GTP, phosphorylates MEK. | Sterile Vul (maternal rescue of rod-like lethal), etc. | Han et al., 1993; Hsu et al., 2002 |
| LIP-1 | Lateral signal Induced Phosphatase | MKP1 | MAPK phosphatase | Partial sterile. Muv in combination with gap-1, others. | Berset et al., 2001 |
| LRP-1 | Low-density Lipoprotein Receptor-related Protein | LRP1 | Lipoprotein Receptor-related protein. Required for EGL-17 secretion. | Abnormal sex myoblast positions. | Kamikura and Cooper, 2003 |
| LST-1 | Lateral Signaling Target | - | Novel MPK-1 binding protein | WT. Muv in combination with gap-1. | Yoo et al., 2004 |
| LST-2 | Lateral Signaling Target | Zinc finger FYVE domain containing protein 28 | FYVE domain protein | WT. Muv in combination with gap-1. | Yoo et al., 2004 |
| LST-3 | Lateral Signaling Target | CCAR1 | SAF-A/B, Acinus and PIAS domain protein | WT. Muv in combination with gap-1. | Yoo et al., 2004 |
| LST-4 | Lateral Signaling Target | NM_153271 | Sorting nexin, may promote LET-23 degradation | WT. Muv in combination with gap-1. | Yoo et al., 2004 |
| MEK-2 | Map kinase kinase or Erk Kinase | MEK1, 2 | Dual specificity kinase, phosphorylates ERK | Sterile Vul (maternal rescue of rod-like lethal), etc. | Kornfeld et al., 1995a; Wu et al, 1995 |
| MPK-1/ SUR-1 | MaP Kinase/ SUppressor of Ras | ERK1, 2 | Serine/Threonine kinase | Sterile Vul (maternal rescue of rod-like lethal), etc. | Lackner and Kim, 1998; Lackner et al., 1994; Wu and Han, 1994 |
| PAR-1 | Abnormal embryonic PARtitioning of cytoplasm | MARK2/C- TAK1 | Serine/Threonine kinase | Weak Muv. Reverses sur-6 suppressor phenotype. | Kao et al., 2004; Yoder et al., 2004 |
| PTP-2 | Protein Tyrosine Phosphatase | Shp-2 | Tyrosine phosphatase | Sterile and maternal-effect lethal. Suppressed by let-60 ras(gf). Suppresses clr-1. | Gutch et al., 1998; Schutzman et al., 2001 |
| ROM-1 | RhOMboid-related | RHBDL2 | 7-pass transmemb- rane serine protease | WT. Suppresses let-60 ras(gf) Muv. | Dutt et al., 2004 |
| SEM-4 | SEx Muscle abnormality | Sal1 | C2H2 zinc finger transcription factor | Partial Vul. | Grant et al., 2000 |
| SEM-5 | SEx Muscle abnormality | Grb2 | RTK-binding adaptor, | Rod-like larval lethal, Vul, etc. | Clark et al., 1992 |
| SLI-1 | Suppressor of LIneage defect | Cbl | E3 ubiquitin ligase, Involved in LET-23 endocytosis/ degradation | WT. Muv in combination with unc-101, others. | Jongeward et al., 1995; Yoon et al., 1995 |
| SOC-1 | Suppressor Of Clr-1 | Gab1 | RTK-binding adaptor, Promotes EGL-15 signaling | Scrawny. Suppresses clr-1 lethality. | Schutzman et al., 2001 |
| SOS-1/ LET-341 | Son Of Sevenless/ LEThal | Sos-1 | Guanine Nucleotide Exchange Factor | Rod-like larval lethal, Vul, etc. | Chang et al., 2000 |
| SRA-13 | Serpentine Receptor, A class | - | G-protein coupled receptor | Increased chemotaxis. Suppresses let-60(dn) Vul. | Battu et al., 2003 |
| SUR-2 | SUppressor of Ras | Sur-2 | Mediator subunit | Vul. | Singh and Han, 1995 |
| SUR-5 | SUppressor of Ras | NM_023928 | Acetyl coenzyme A synthetase | WT. Suppresses let-60(dn) Vul. | Gu et al., 1998 |
| SUR-6 | SUppressor of Ras | PPP2R2A | PR55/B Regulatory subunit of Protein Phosphatase 2A | Suppresses let-60 ras(gf) Muv. Enhances Vul and rod-like lethal phenotypes of lin-45 hypomorphs, others. | Kao et al., 2004; Sieburth et al., 1999 |
| SUR-7 | SUppressor of Ras | - | Zinc transporter | Suppresses let-60 ras(gf) Muv. | Yoder et al., 2004 |
| SUR-8/ SOC-2 | SUppressor of Ras, Suppressor Of Clr-1 | Sur-8 | Ras-binding Leucine-rich repeat protein | Scrawny. Suppresses let-60 ras(gf) Muv and clr-1 lethality. Enhances Vul and rod-like lethal phenotypes of lin-45 hypomorphs, others. | Selfors et al., 1998; Sieburth et al., 1998 |
| UNC-101 | UNCoordinated | AP-47 | Clathrin adaptor subunit | WT. Muv in combination with sli-1, others. | Lee et al., 1994 |
A general model for the core C. elegans RTK/Ras/MAPK signaling pathway (and various tissue-specific variations) is shown in Figure 1 (Moghal and Sternberg, 2003; Schlessinger, 2000). Upon growth factor binding, an RTK such as LET-23 or EGL-15 dimerizes and autophosphorylates its C-terminal region. The resulting phospho-tyrosine residues serve as docking sites for adaptor proteins such as SEM-5 (Grb2) or SOC-1 (similar to Gab1). These adaptors recruit the Guanine Nucleotide Exchange Factor SOS-1 to activate the small GTPase LET-60 Ras. LET-60-GTP then binds to LIN-45 Raf and promotes its stable association with the plasma membrane and/or endomembranes, where other events then activate LIN-45 kinase activity (Chong et al., 2003). The scaffold protein KSR may assist in LIN-45 activation, but also promotes further signal transmission by bringing together different components of the MAPK cascade (Morrison and Davis, 2003). LIN-45 phosphorylates and activates MEK-2, MEK-2 phosphorylates and activates MPK-1, and MPK-1 then phosphorylates and either activates or inactivates various target proteins. In many cases MPK-1 may move into the nucleus to phosphorylate transcription factors such as the Ets domain protein LIN-1, thus leading to changes in gene expression.
This model draws on a large body of data from multiple systems. C. elegans genetics has been most useful for identifying the genes involved in particular signaling events and determining their order of action and cellular focus. In some cases, physical interactions and/or phosphorylation events have been demonstrated for the worm proteins (e.g., Jacobs et al., 1999; Sieburth et al., 1998; Wu et al., 1995), but in many cases such interactions are inferred based on biochemical studies of related proteins in vertebrate cells. Because the core Ras pathway (and much of its regulation) appears highly conserved between C. elegans and vertebrates (Table 1), current models draw on the combined data from these different systems.
Ras signaling is not required for cellular viability in C. elegans (Yochem et al., 1997), but it is required for organismal viability and for many different developmental processes. Because of the widespread roles of Ras signaling during development, mutations affecting the Ras pathway can cause many different pleiotropic defects (Figure 2).
|
Figure 2. let-60 ras mutant phenotypes. Wild-type phenotypes are shown on the left, Ras pathway mutant phenotypes on the right. See text for details.
Lethality: Ras signaling promotes the excretory duct cell fate, and mosaic analysis suggested that loss of this one specific cell can account for the zygotic lethality of let-60 null mutants (Yochem et al., 1997). The excretory duct cell is required for osmoregulation (Nelson and Riddle, 1984). let-60 loss-of-function mutants, which lack the excretory duct cell, die as “rod-like” larvae with a fluid-filled appearance (Figure 2). let-60 gain-of-function mutants sometimes have two excretory duct cells (Yochem et al., 1997). Ras signaling may have other (maternally-rescued) essential roles in addition to excretory duct formation, since mutations in let-23/EGFR, egl-15/FGFR and ptp-2/SHP-2 cause distinct “scrawny” and lethal defects that are rescued by constitutive forms of LET-60 Ras (DeVore et al., 1995; Gutch et al., 1998; Koga and Ohshima, 1995).
Hyperactivation of egl-15/FGFR or let-60/Ras can also lead to lethality with a “Clear”, fluid-filled appearance (Kokel et al., 1998; Schutzman et al., 2001). Mosaic analysis suggests that the Clear phenotype is caused by hypodermal defects (Huang and Stern, 2004).
Vulval defects: Ras signaling cooperates with Notch and Wnt signaling to promote hermaphrodite vulval development (see Vulval development). let-60 loss-of-function mutants lack a vulva (Vulvaless or Vul phenotype; Figure 2) whereas let-60 gain-of-function mutants have extra vulval tissue (Multivulva or Muv phenotype).
Uterine defects: Ras signaling promotes the uterine uv1 fate, which is important for establishing a proper vulval-uterine connection (Chang et al., 1999). In the absence of uv1, hermaphrodites cannot lay eggs.
P11 and P12 ectoblast defects: Ras signaling cooperates with Wnt signaling to promote the P12 ectoblast cell fate (Fixsen et al., 1985; Jiang and Sternberg, 1998). The P11 and P12 ectoblasts are neighboring cells that give rise to slightly different types of hypodermal and neuronal descendants (Sulston and Horvitz, 1977; Hermaphrodite cell fate specification). Reduced Ras signaling causes P12-> P11 cell fate transformations (Figure 2), while increased signaling causes P11-> P12 fate transformations.
Male spicule defects: Ras signaling promotes male spicule fates (Chamberlin and Sternberg, 1994; Male development). Reduced Ras signaling causes spicule defects and prevents males from mating.
Sex myoblast migration defects: Ras signaling helps to specify the proper endpoint of sex myoblast migration (Sundaram et al., 1996). In let-60 loss-of-function mutants, sex myoblasts adopt a broadened range of final positions. This defect can affect egg-laying.
Axon guidance defects: Ras signaling controls the paths of certain ventral cord neurons relative to the ventral midline (Bulow et al., 2004). Whereas in wild-type these neurons extend axons along one side of the midline, in let-60 loss-of-function mutants the neurons wander across the midline.
Sterility due to pachytene exit (Pex) defects: Ras signaling is required for progression through the pachytene stage of meiosis (Church et al., 1995; Somatic sex determination). let-60 loss-of-function mutants are sterile because germ cells arrest in pachytene (Figure 2).
Olfaction defects: Ras signaling is required for sensitivity to volatile attractants (Hirotsu et al., 2000). Whereas wild-type C. elegans will chemotax toward volatile attractants such as isoamylalcohol and diacetyl, let-60 loss-of-function mutants fail to chemotax towards such attractants.
Resistance to Microbacterium nematophilum-induced swelling: ksr-1, lin-45, mek-2 and mpk-1 are required for the swelling response to infection by M. nematophilum (Nicholas and Hodgkin, 2004). Interestingly, let-60 does not appear to be important for this response, suggesting that a bacterial toxin may directly activate the Raf/MEK/ERK cascade.
The vast majority of studies on RTK/Ras/MAPK signaling have focused on the role of the Ras pathway in promoting vulval development (see Vulval development), and many known components of the pathway have been identified through forward genetic screens for mutants with Vul or Muv mutant phenotypes (Ferguson and Horvitz, 1985). Some components of the pathway have been identified based on other mutant phenotypes such as sex myoblast migration defects (Stern and Horvitz, 1991) or germline meiosis defects (Church et al., 1995; Ohmachi et al., 2002). Some have been identified through reverse genetic approaches (Dutt et al., 2004; Gutch et al., 1998; Kamikura and Cooper, 2003; Yoo et al., 2004). Finally, some core Ras pathway components as well as a large set of other genes that influence Ras signaling have been identified through genetic suppressor or enhancer screens (Sternberg and Han, 1998). Many modifier genes have essentially wild-type null phenotypes (Table 1), indicating a significant and surprising amount of redundancy amongst Ras pathway regulators.
One especially productive modifier screen involved looking for suppressors of the let-60 gain-of-function Muv phenotype. This screen identified alleles of the downstream kinase genes lin-45/Raf (Hsu et al., 2002), mek-2/MEK (Kornfeld et al., 1995; Wu et al., 1995) and mpk-1/ERK (Lackner et al., 1994; Wu and Han, 1994), revealing that LET-60 Ras signals through the MAPK cascade. This screen also identified numerous positive regulators of this cascade such as cdf-1 (Jakubowski and Kornfeld, 1999), ksr-1 (Kornfeld et al., 1995; Sundaram and Han, 1995), sur-6 (Sieburth et al., 1999), sur-7 (Yoder et al., 2004), and sur-8 (Sieburth et al., 1998; see below), as well as a key downstream factor, sur-2 (Singh and Han, 1995; see below). Another very productive type of screen has been to look for enhancers of lethal or vulval defects in the background of a very mildly affected or “phenotype-less” mutant. For example, screens for enhancers of lin-45 lethality identified the downstream factors eor-1 and eor-2 (Howard and Sundaram, 2002; Rocheleau et al., 2002), while screens for enhancers of the gap-1 Muv phenotype identified the MAPK phosphatase lip-1 (Berset et al., 2001). These types of genetic modifier screens have allowed the identification of genes with very subtle null phenotypes.
The power of genetic screening approaches in C. elegans is reflected in the large number of conserved genes whose involvement in RTK/Ras/MAPK signaling was first discovered in the worm; these include ark-1 (Hopper et al., 2000), cdf-1/ZnT1 (Jakubowski and Kornfeld, 1999), ksr-1 (Kornfeld et al., 1995; Sundaram and Han, 1995), lin-2/CASK, lin-7 and lin-10/Mint (Ferguson and Horvitz, 1985), sem-5/Grb2 (Clark et al., 1992), sli-1/Cbl (Jongeward et al., 1995), sur-2 (Singh and Han, 1995), sur-5 (Gu et al., 1998), sur-7 (Yoder et al., 2004), and sur-8/soc-2 (Selfors et al., 1998; Sieburth et al., 1998).
The C. elegans genome contains twenty-eight predicted RTKs (Popovici et al., 1999), only a few of which have been characterized mutationally (see Genomic overview of protein kinases). Of these characterized RTKs, only the EGF receptor LET-23 and the FGF receptor EGL-15 are known to signal positively through Ras/MAPK (see below). The ephrin receptor VAB-1 negatively regulates MAPK activation during oocyte maturation (Miller et al., 2003). Notably, the insulin-like RTK DAF-2 does not appear to signal through Ras/MAPK (G. Ruvkun, personal communication), but instead signals through a PI3-kinase/Akt pathway (see Signaling in the immune response).
Signaling by the LET-23 and EGL-15 RTKs can account for many but not all LET-60 Ras-dependent developmental events. The EGF-related ligand LIN-3 signals through LET-23/EGFR and LET-60 to control specification of hermaphrodite vulval and uterine fates, male spicule cell fates, the P12 ectoblast cell fate, and most likely the excretory duct cell fate (Aroian and Sternberg, 1991; Chang et al., 1999; Figure 1A). The FGF-related ligand EGL-17 signals through EGL-15/FGFR and LET-60 to control sex myoblast migration (Figure 1B), while the FGF-related ligand LET-756 signals through EGL-15 and LET-60 to promote axon guidance and viability (Bulow et al., 2004; Burdine et al., 1997; DeVore et al., 1995; Goodman et al., 2003; Roubin et al., 1999; Sundaram et al.,1996). In addition, LET-60 acts independently of any known RTK to control olfaction (Hirotsu et al., 2000) and the progression of germline meiosis (Church et al., 1995; Figure 1C). Furthermore, LIN-45, MEK-2 and MPK-1 act independently of known RTKs or LET-60 to mediate responses to Microbacterium nematophilum infection (Nicholas and Hodgkin, 2004).
In some cases the LET-23 and EGL-15 RTKs can also act through LET-60 Ras-independent pathways. LIN-3 and LET-23 control oocyte maturation via PLC gamma and an inositol polyphosphate signaling pathway (Bui and Sternberg, 2002; Clandinin et al., 1998; Yin et al., 2004). EGL-17 and EGL-15 may act through additional, unknown pathways to control sex myoblast migration and osmoregulation (DeVore et al., 1995; Schutzman et al., 2001; Sundaram et al., 1996).
Reverse genetic approaches have identified several factors important for ligand processing and secretion. The ligand LIN-3 exists in both trans-membrane (locally-acting) and diffusible forms (Hill and Sternberg, 1992; Thomas et al., 1990); generation of the diffusible form in vulval cells appears to require cleavage by the Rhomboid ortholog ROM-1 (Dutt et al., 2004). The ligand EGL-17 is secreted via a mechanism that requires the lipoprotein receptor-related proteins LRP-1 and LRP-2 and the Disabled-related adaptor DAB-1 (Kamikura and Cooper, 2003).
Factors that regulate LET-23 RTK trafficking and localization are important modulators of signaling strength. A complex consisting of three PDZ-domain proteins, LIN-2, LIN-7 and LIN-10, positively regulates signaling by localizing LET-23 to the basolateral membrane of vulval precursor cells, adjacent to the source of LIN-3 ligand (Kaech et al., 1998; Simske et al., 1996). The E3 ubiquitin ligase SLI-1 (Cbl; Jongeward et al., 1995; Yoon et al., 1995), the clathrin adaptors UNC-101 (Lee et al., 1994) and DPY-23 (Yoo et al., 2004), and the sorting nexin LST-4 (Yoo et al., 2004) all negatively regulate signaling, probably by promoting LET-23 endocytosis and/or degradation.
RTKs transduce signals by autophosphorylation and subsequent binding to phospho-tyrosine binding adaptor proteins (Schlessinger, 2000). LET-23 seems to act primarily through the adaptor SEM-5 (Clark et al., 1992), while EGL-15 acts through both SEM-5 and the adaptor SOC-1 (Schutzman et al., 2001). Factors that regulate RTK phosphorylation status could obviously modulate signaling strength. For example, the receptor tyrosine phosphatase CLR-1 negatively regulates EGL-15 RTK activity, most likely through direct dephosphorylation of key tyrosine residues (Kokel et al., 1998). On the other hand, the cytosolic tyrosine phosphatase PTP-2 (Shp-2) positively regulates LET-23 and EGL-15 RTK signaling (Gutch et al., 1998; Schutzman et al., 2001), and the tyrosine kinase ARK-1 (Ack-related) negatively regulates LET-23 RTK signaling (Hopper et al., 2000). The substrates and mechanisms of action of PTP-2 and ARK-1 are still unclear, but these proteins can bind to the adaptors SOC-1 and SEM-5, respectively, and appear to act at a step immediately downstream of the RTKs.
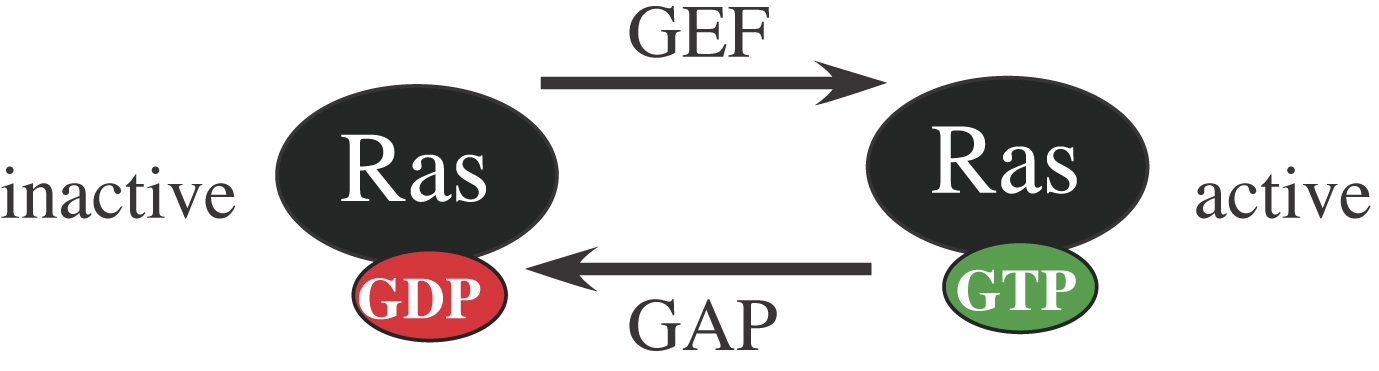
As is the case for other small GTPases (see Small GTPases), the activity of LET-60 Ras is controlled by Guanine Nucleotide Exchange Factors (GEFs), which activate Ras by stimulating conversion of Ras-GDP to Ras-GTP, and by GTPase activating proteins (GAPs), which inactivate Ras by stimulating conversion of Ras-GTP to Ras-GDP (Figure 3). The GEF SOS-1 appears necessary for most Ras-mediated developmental events (Chang et al., 2000). The GAPs GAP-1 and GAP-2 negatively regulate Ras signaling during vulval development and excretory duct development, respectively (Hajnal et al., 1997; Hayashizaki et al., 1998). Another negative regulator of Ras is SUR-5, a protein of unknown function that resembles acetyl coenzyme A synthetases (Gu et al., 1998).
|
Figure 3. Regulation of Ras proteins. Ras GTPase regulation has been extensively studied in other systems (Malumbres and Barbacid, 2002). Ras-GDP is inactive, whereas Ras-GTP is active and can bind to effectors such as Raf. Guanine nucleotide exchange factors (GEFs) such as SOS-1 positively regulate Ras by promoting GDP dissociation. GTPase activating proteins (GAPs) negatively regulate Ras by stimulating Ras’ intrinsic GTP hydrolyzing activity. Gain-of-function (gf) mutations lock Ras in the active, GTP-bound state; the let-60(gf) allele n1046 (G13E) has been widely used for genetic analyses (Beitel et al., 1990; Han and Sternberg, 1990). Dominant-negative (dn) mutations lock Ras in the inactive, GDP-bound state, causing it to bind stably to and titrate out GEFs; a variety of let-60(dn) mutations have been described (Han and Sternberg, 1991).
Genetic modifier screens have identified a large set of genes that promote or inhibit signaling through the MAPK cascade, but that are not individually required for normal development in most tissues. In general, the roles of these genes can only be detected as suppressor or enhancer effects in appropriate double mutant combinations. Raf/MEK/ERK signaling is apparently subject to many different levels of regulation that individually have modest effects on signaling strength.
Positively-acting gene products in this category include so-called “scaffold” proteins - CNK-1 (Rocheleau et al., 2005), KSR-1 (Kornfeld et al., 1995; Sundaram and Han, 1995), KSR-2 (Ohmachi et al., 2002), and SUR-8/SOC-2 (Selfors et al., 1998; Sieburth et al., 1998) - that bind to one or more core components of the signaling cascade and are thought to bring different components together at the right place and time (Morrison and Davis, 2003). KSR-1 and KSR-2 are individually required for some developmental processes, but redundantly required for many others (Ohmachi et al., 2002; Figure 1). Also in this category is SUR-6, a PR55/B-type regulatory subunit of Protein Phosphatase 2A (Sieburth et al., 1999), which has been suggested to dephosphorylate and activate both Raf and KSR (Kao et al., 2003; Ory et al., 2003). Finally, the zinc transporter proteins CDF-1 and SUR-7 also positively regulate signaling, suggesting that high levels of intracellular zinc inhibit Ras signaling (Bruinsma et al., 2002; Yoder et al., 2004).
Negatively acting gene products in this category include the G-protein coupled receptor SRA-13 and its Gα target GPA-5, which may modulate Ras signaling in response to environmental conditions such as food availability (Battu et al., 2003). Also in this category are the kinase PAR-1 (which is thought to modulate KSR localization; Kao et al., 2003; Muller et al., 2001; Yoder et al., 2004), the MPK-1-binding protein LST-1 (Yoo et al., 2004), and the MAP kinase phosphatase LIP-1 (Berset et al., 2001).
It is important to note that some regulatory proteins may influence Raf/MEK/ERK signaling indirectly, by affecting neighboring cells. For example, the Gαq protein EGL-30 and the voltage-gated calcium channel EGL-19 promote vulval induction, but function in neurons and muscle, respectively (Moghal et al., 2003). Similarly, SRA-13 and GPA-5 influence vulval induction but it is unknown whether these genes function in vulval cells or in neurons (Battu et al., 2003). Also, the zinc transporter CDF-1 can influence vulval induction when expressed in either the vulva or the intestine (Bruinsma et al., 2002). How tissues such as the intestine, neurons and muscle can influence Raf/MEK/ERK activity in vulval cells is still unclear.
No single downstream target of MPK-1 can account for all of the effects of RTK/Ras/MAPK signaling. Rather, different tissues seem to require different subsets of potential targets, and the availability of certain targets may control tissue-specific responses.
One widely important MPK-1 target is the Ets domain transcription factor LIN-1 (Beitel et al., 1995; Figure 1A). LIN-1 antagonizes Ras signaling and appears to be negatively regulated by MPK-1 phosphorylation (Jacobs et al., 1998). However, some evidence suggests an additional positive role in Ras signaling (Howard and Sundaram, 2002). Like other Ets domain factors (Yordy and Muise-Helmericks, 2000), LIN-1 may have both transcriptional repressor and transcriptional activator functions.
Four widely important positive factors are the Mediator subunit SUR-2, the BTB/Zinc finger protein EOR-1, and the novel nuclear proteins LIN-25 and EOR-2 (Howard and Sundaram, 2002; Singh and Han, 1995; Tuck and Greenwald, 1995; Figure 1A). None of these proteins are known to be direct targets of MPK-1, but their functions are important for downstream cellular responses. SUR-2 is a conserved component of the Mediator complex, which links certain sequence-specific DNA binding proteins (such as Ets proteins) to the general RNA Polymerase II transcriptional machinery (Boyer et al., 1999; Stevens et al., 2002). SUR-2 and LIN-25 appear to function together, and they have strong effects on vulval development and weaker effects on excretory duct and P12 development (Nilsson et al., 1998; Nilsson et al., 2000; Singh and Han, 1995; Tuck and Greenwald, 1995). EOR-1 is related to known transcriptional activators and repressors (Barna et al., 2002; Collins et al., 2001). EOR-1 and EOR-2 appear to function together, and they have moderate effects on excretory duct and P12 development and weaker effects on vulval development (Howard and Sundaram, 2002). EOR-1 and EOR-2 appear to act redundantly with SUR-2 and LIN-25 (Howard and Sundaram, 2002).
Several transcription factors are required for cell-type specific responses to RTK/Ras/MAPK signaling, and are candidate MPK-1 targets. These include the forkhead transcription factor LIN-31 (Miller et al., 1993; Tan et al., 1998), which both promotes and inhibits vulval development, the Hox protein LIN-39 (Clark et al., 1993; Eisenmann et al., 1998; Maloof and Kenyon, 1998), the zinc-finger protein SEM-4 (Grant et al., 2000) and the GATA factors EGL-18 and ELT-6 (Koh et al., 2004; Koh et al., 2002), which promote vulval development, and the Hox protein EGL-5 (Chisholm, 1991; Jiang and Sternberg, 1998), which promotes P12 development. Ectopic expression of LIN-31 or LIN-39 can cause other tissues to adopt vulval-like characteristics in response to RTK/Ras/MAPK signaling, suggesting that the presence of these distinct transcription factors may be a key determinant of tissue-specific responses to Ras signaling (Maloof and Kenyon, 1998; Tan et al., 1998).
Not all targets of MPK-1 need be transcription factors. For example, none of the widely important transcriptional regulators seem to be involved in controlling germline meiosis or sex myoblast migration (Church et al., 1995; Sundaram et al., 1996; R. Howard and M. Sundaram, unpublished observations; Figure 1B and C). It is likely that MPK-1 targets in these tissues include factors more directly involved in meiosis and motility.
The RTK/Ras/MAPK pathway often interacts with other signaling pathways to control cell fates. For example, the Ras pathway cooperates with a Wnt pathway (see Wnt signaling) to specify P12 fates and vulval fates, possibly by convergent upregulation of common targets such as Hox genes (Eisenmann et al., 1998; Gleason et al., 2002). During vulval development (see Vulval development), the Ras pathway also acts sequentially with a Notch pathway (see LIN-12/Notch signaling in C. elegans) to induce the proper pattern of vulval fates (Simske and Kim, 1995; Sundaram, 2004). Ras signaling affects Notch signaling in at least two ways. First, Ras stimulates LIN-12/Notch endocytosis to downregulate Notch signaling in the same cell (Shaye and Greenwald, 2002). Second, Ras stimulates the transcription of Notch ligand genes to upregulate Notch signaling in adjacent cells (Chen and Greenwald, 2004). Notch signaling also antagonizes Ras signaling by stimulating the transcription of various negative regulators such as lip-1 and lst-1-4 (Berset et al., 2001; Yoo et al., 2004).
RTK/Ras/MAPK signaling in both P12 and the vulva is antagonized by LIN-35 (Rb), EFL-1 (E2F) and other “Synthetic Multivulva” (SynMuv) gene products (Ceol and Horvitz, 2001; Fay and Han, 2000; Jiang and Sternberg, 1998; Lu and Horvitz, 1998). There are three classes of SynMuv genes, A, B and C, that function redundantly (Ceol and Horvitz, 2004; Ferguson and Horvitz, 1989). Many of these genes encode nuclear proteins with apparent roles in chromatin remodeling and transcriptional regulation (Fay and Han, 2000). Some SynMuv genes seem to function cell autonomously and could antagonize Ras signaling at the level of downstream transcriptional output (Thomas and Horvitz, 1999). However, other SynMuv genes (including lin-35/Rb) appear to function cell non-autonomously in the syncytial hypodermis hyp7, suggesting that hyp7-derived signals can influence LET-23 RTK activity (Herman and Hedgecock, 1990; Myers and Greenwald, 2005).
Although the basic framework of the RTK/Ras/MAPK pathway is well characterized, the pathway is subject to complex regulation that we are only just beginning to understand. A most surprising finding in C. elegans has been the large number of regulatory proteins that have only modest individual effects on signaling. Furthermore, some regulatory proteins (such as LIN-2/LIN-7/LIN-10) have very cell-type specific effects. Downstream targets of MPK-1 also appear to be cell-type specific, and likely control how a particular cell responds to the same basic signaling pathway. Although vulval development has been and will continue to be a powerful model for studying RTK/Ras/MAPK signaling, ongoing studies of other Ras-mediated developmental processes will surely reveal new and different types of regulatory mechanisms and targets.
My laboratory’s work on RTK/Ras/MAPK signaling is supported by NIH grants GM58540 and CA87512.
Aroian, R.V., Koga, M., Mendel, J.E., Ohshima, Y., and Sternberg, P.W. (1990). The let-23 gene necessary for Caenorhabditis elegans vulval induction encodes a tyrosine kinase of the EGF receptor subfamily [see comments]. Nature 348, 693–699. Abstract Article
Aroian, R.V., and Sternberg, P.W. (1991). Multiple functions of let-23, a Caenorhabditis elegans receptor tyrosine kinase gene required for vulval induction. Genetics 128, 251–267. Abstract
Barna, M., Merghoub, T., Costoya, J.A., Ruggero, D., Branford, M., Bergia, A., Samori, B., and Pandolfi, P.P. (2002). Plzf mediates transcriptional repression of HoxD gene expression through chromatin remodeling. Dev. Cell 3, 499–510. Abstract Article
Battu, G., Hoier, E.F., and Hajnal, A. (2003). The C. elegans G-protein-coupled receptor SRA-13 inhibits RAS/MAPK signalling during olfaction and vulval development. Development 130, 2567–2577. Abstract Article
Beitel, G.J., Clark, S.G., and Horvitz, H.R. (1990). Caenorhabditis elegans ras gene let-60 acts as a switch in the pathway of vulval induction. Nature 348, 503–509. Abstract Article
Beitel, G.J., Tuck, S., Greenwald, I., and Horvitz, H.R. (1995). The Caenorhabditis elegans gene lin-1 encodes an ETS-domain protein and defines a branch in the vulval induction pathway. Genes Dev. 9, 3149–3162. Abstract
Berset, T., Hoier, E.F., Battu, G., Canevascini, S., and Hajnal, A. (2001). Notch inhibition of RAS signaling through MAP kinase phosphatase LIP-1 during C. elegans vulval development. Science 291, 1055–1058. Abstract Article
Boyer, T.G., Martin, M.E., Lees, E., Ricciardi, R.P., and Berk, A.J. (1999). Mammalian Srb/Mediator complex is targeted by adenovirus E1A protein [see comments]. Nature 399, 276–279. Abstract Article
Bruinsma, J.J., Jirakulaporn, T., Muslin, A.J., and Kornfeld, K. (2002). Zinc ions and cation diffusion facilitator proteins regulate Ras-mediated signaling. Dev. Cell 2, 567–578. Abstract Article
Bui, Y.K., and Sternberg, P.W. (2002). Caenorhabditis elegans inositol 5-phosphatase homolog negatively regulates inositol 1,4,5-triphosphate signaling in ovulation. Mol. Biol. Cell 13, 1641–1651. Article
Bulow, H.E., Boulin, T., and Hobert, O. (2004). Differential functions of the C. elegans FGF receptor in axon outgrowth and maintenance of axon position. Neuron 42, 367–374. Abstract Article
Burdine, R.D., Chen, E.B., Kwok, S.F., and Stern, M.J. (1997). egl-17 encodes an invertebrate fibroblast growth factor family member required specifically for sex myoblast migration in Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 94, 2433–2437. Abstract Article
Ceol, C.J., and Horvitz, H.R. (2001). dpl-1 DP and efl-1 E2F Act with lin-35 Rb to Antagonize Ras Signaling in C. elegans Vulval Development. Mol. Cell 7, 461–473. Abstract Article
Ceol, C.J., and Horvitz, H.R. (2004). A new class of C. elegans SynMuv genes implicates a Tip60/NuA4-like HAT complex as a negative regulator of Ras signaling. Dev. Cell 6, 563–576. Abstract Article
Chamberlin, H.M., and Sternberg, P.W. (1994). The lin-3/let-23 pathway mediates inductive signalling during male spicule development in Caenorhabditis elegans. Development 120, 2713–2721. Abstract
Chang, C., Hopper, N.A., and Sternberg, P.W. (2000). Caenorhabditis elegans SOS-1 is necessary for multiple RAS-mediated developmental signals. EMBO J. 19, 3283–3294. Abstract Article
Chang, C., Newman, A.P., and Sternberg, P.W. (1999). Reciprocal EGF signaling back to the uterus from the induced C. elegans vulva coordinates morphogenesis of epithelia. Curr. Biol. 9, 237–246. Abstract Article
Chen, N., and Greenwald, I. (2004). The lateral signal for LIN-12/Notch in C. elegans vulval development comprises redundant secreted and transmembrane DSL proteins. Dev. Cell 6, 183–192. Abstract Article
Chisholm, A. (1991). Control of cell fate in the tail region of C. elegans by the gene egl-5. Development 111, 921–932. Abstract
Chong, H., Vikis, H.G., and Guan, K.-L. (2003). Mechanisms of regulating the Raf kinase family. Cell. Signal 15, 463–469. Abstract Article
Church, D., Guan, K.L., and Lambie, E.J. (1995). Three genes of the MAP kinase cascade, mek-2, mpk-1/sur-1 and let-60 ras, are required for meiotic cell cycle progression in Caenorhabditis elegans. Development 121, 2525–2535. Abstract
Clandinin, T.R., DeModena, J.A., and Sternberg, P.W. (1998). Inositol trisphosphate mediates a RAS-independent response to LET-23 receptor tyrosine kinase activation in C. elegans. Cell 92, 523–533. Abstract Article
Clark, S.G., Chisholm, A.D., and Horvitz, H.R. (1993). Control of cell fates in the central body region of C. elegans by the homeobox gene lin-39. Cell 74, 43–55. Abstract Article
Clark, S.G., Stern, M.J., and Horvitz, H.R. (1992). C. elegans cell-signaling gene sem-5 encodes a protein with SH2 and SH3 domains. Nature 356, 340–344. Abstract Article
Collins, T., Stone, J.R., and Williams, A.J. (2001). All in the family: the BTB/POZ, KRAB and SCAN domains. Mol. Cell. Biol. 21, 3609–3615. Abstract Article
DeVore, D.L., Horvitz, H.R., and Stern, M.J. (1995). An FGF receptor signaling pathway is required for the normal cell migrations of the sex myoblasts in C. elegans hermaphrodites. Cell 83, 611–620. Abstract Article
Dutt, A., Canevascini, S., Froehli-Hoier, E., and Hajnal, A. (2004). EGF signal propagation during C. elegans vulval development mediated by ROM-1 Rhomboid. PLoS Biol. 2, 0001–0016. Abstract Article
Eisenmann, D.M., Maloof, J.N., Simske, J.S., Kenyon, C., and Kim, S.K. (1998). The beta-catenin homolog BAR-1 and LET-60 Ras coordinately regulate the Hox gene lin-39 during Caenorhabditis elegans vulval development. Development 125, 3667–3680. Abstract
Fay, D.S., and Han, M. (2000). The synthetic multivulval genes of C. elegans: functional redundancy, Ras-antagonism, and cell fate determination. Genesis: J. Genet. Dev. 26, 279–284. Abstract
Ferguson, E.L., and Horvitz, H.R. (1985). Identification and characterization of 22 genes that affect the vulval cell lineages of the nematode Caenorhabditis elegans. Genetics 110, 17–72. Abstract
Ferguson, E.L., and Horvitz, H.R. (1989). The multivulva phenotype of certain Caenorhabditis elegans mutants results from defects in two functionally redundant pathways. Genetics 123, 109–121. Abstract
Fixsen, W., Sternberg, P., Ellis, H., and Horvitz, R. (1985). Genes that affect cell fates during the development of Caenorhabditis elegans. Cold Spring Harb. Symp. Quant. Biol. 50, 99–104. Abstract
Gleason, J.E., Korswagen, H.C., and Eisenmann, D.M. (2002). Activation of Wnt signaling bypasses the requirement for RTK/Ras signaling during C. elegans vulval induction. Genes Dev. 16, 1281–1290. Abstract Article
Goodman, S.J., Branda, C.S., Robinson, M.K., Burdine, R.D., and Stern, M.J. (2003). Alternative splicing affecting a novel domain in the C. elegans EGL-15 FGF receptor confers functional specificity. Development 130, 3757–3766. Abstract Article
Grant, K., Hanna-Rose, W., and Han, M. (2000). sem-4 promotes vulval cell-fate determination in Caenorhabditis elegans through regulation of lin-39 Hox. Dev. Biol. 224, 496–506. Abstract Article
Gu, T., Orita, S., and Han, M. (1998). Caenorhabditis elegans SUR-5, a novel but conserved protein, negatively regulates LET-60 Ras activity during vulval induction. Mol. Cell Biol. 18, 4556–4564. Abstract
Gutch, M.J., Flint, A.J., Keller, J., Tonks, N.K., and Hengartner, M.O. (1998). The Caenorhabditis elegans SH2 domain-containing protein tyrosine phosphatase PTP-2 participates in signal transduction during oogenesis and vulval development. Genes Dev. 12, 571–585. Abstract
Hajnal, A., Whitfield, C.W., and Kim, S.K. (1997). Inhibition of Caenorhabditis elegans vulval induction by gap-1 and by let-23 receptor tyrosine kinase. Genes Dev. 11, 2715–2728. Abstract
Han, M., Golden, A., Han, Y., and Sternberg, P.W. (1993). C. elegans lin-45 raf gene participates in let-60 ras-stimulated vulval differentiation. Nature 363, 133–140. Abstract Article
Han, M., and Sternberg, P.W. (1990). let-60, a gene that specifies cell fates during C. elegans vulval induction, encodes a ras protein. Cell 63, 921–931. Abstract Article
Han, M., and Sternberg, P.W. (1991). Analysis of dominant-negative mutations of the Caenorhabditis elegans let-60 ras gene. Genes Dev. 5, 2188–2198. Abstract
Hayashizaki, S., Iino, Y., and Yamamoto, M. (1998). Characterization of the C. elegans gap-2 gene encoding a novel Ras-GTPase activating protein and its possible role in larval development. Genes Cells 3, 189–202. Abstract Article
Herman, R.K., and Hedgecock, E.M. (1990). Limitation of the size of the vulval primordium of Caenorhabditis elegans by lin-15 expression in the surrounding hypodermis. Nature 348, 169–171. Abstract Article
Hill, R.J., and Sternberg, P.W. (1992). The gene lin-3 encodes an inductive signal for vulval development in C. elegans [see comments]. Nature 358, 470–476. Abstract Article
Hirotsu, T., Saeki, S., Yamamoto, M., and Iino, Y. (2000). The Ras-MAPK pathway is important for olfaction in Caenorhabditis elegans. Nature 404, 289–293. Abstract Article
Hopper, N.A., Lee, J., and Sternberg, P.W. (2000). ARK-1 inhibits EGFR signaling in C. elegans. Mol. Cell 6, 65–75. Abstract Article
Howard, R.M., and Sundaram, M.V. (2002). C. elegans EOR-1/PLZF and EOR-2 positively regulate Ras and Wnt signaling and function redundantly with LIN-25 and the SUR-2 Mediator complex. Genes Dev. 16, 1815–1827. Abstract Article
Hsu, V., Zobel, C.L., Lambie, E.J., Schedl, T., and Kornfeld, K. (2002). Caenorhabditis elegans lin-45 raf is essential for larval viability, fertility and the induction of vulval cell fates. Genetics 160, 481–492. Abstract
Huang, P., and Stern, M.J. (2004). FGF signaling functions in the hypodermis to regulate fluid balance in C. elegans. Development 131, 2595–2604. Abstract Article
Jacobs, D., Beitel, G.J., Clark, S.G., Horvitz, H.R., and Kornfeld, K. (1998). Gain-of-function mutations in the Caenorhabditis elegans lin-1 ETS gene identify a C-terminal regulatory domain phosphorylated by ERK MAP kinase. Genetics 149, 1809–1822. Abstract
Jacobs, D., Glossip, D., Xing, H., Muslin, A.J., and Kornfeld, K. (1999). Multiple docking sites on substrate proteins form a modular system that mediates recognition by ERK MAP kinase. Genes Dev. 13, 163–175. Abstract Article
Jakubowski, J., and Kornfeld, K. (1999). A local, high-density, single-nucleotide polymorphism map used to clone Caenorhabditis elegans cdf-1. Genetics 153, 743–752. Abstract
Jiang, L.I., and Sternberg, P.W. (1998). Interactions of EGF, Wnt and HOM-C genes specify the P12 neuroectoblast fate in C. elegans. Development 125, 2337–2347. Abstract
Jongeward, G.D., Clandinin, T.R., and Sternberg, P.W. (1995). sli-1, a negative regulator of let-23-mediated signaling in C. elegans. Genetics 139, 1553–1566. Abstract
Kaech, S.M., Whitfield, C.W., and Kim, S. (1998). The LIN-2/LIN-7/LIN-10 complex mediates basolateral membrane localization of the C. elegans EFG receptor LET-23 in vulval epithelial cells. Cell 94, 761–771. Abstract Article
Kamikura, D.M., and Cooper, J.A. (2003). Lipoprotein receptors and a disabled family cytoplasmic adaptor protein regulate EGL-17/FGF export in C. elegans. Genes Dev. 17, 2798–2811. Abstract Article
Kao, G., Tuck, S., Baillie, D., and Sundaram, M.V. (2003). C. elegans SUR-6/PR55 cooperates with LET-92/protein phosphatase 2A and promotes Raf activity independently of inhibitory Akt phosphorylation sites. Development 131, 755–765. Abstract Article
Koga, M., and Ohshima, Y. (1995). Mosaic analysis of the let-23 gene function in vulval induction of Caenorhabditis elegans. Development 121, 2655–2666. Abstract
Koh, K., Bernstein, Y., and Sundaram, M.V. (2004). The nT1 translocation separates vulval regulatory elements from the egl-18 and elt-6 GATA factor genes. Dev. Biol. 267, 252–263. Abstract Article
Koh, K., Peyrot, S.M., Wood, C.G., Wagmaister, J.A., Maduro, M.F., Eisenmann, D.M., and Rothman, J.H. (2002). Cell fates and fusion in the C. elegans vulval primordium are regulated by the EGL-18 and ELT-6 GATA factors -- apparent direct targets of the LIN-39 Hox protein. Development 129, 5171–5180. Abstract
Kokel, M., Borland, C.Z., DeLong, L., Horvitz, H.R., and Stern, M.J. (1998). clr-1 encodes a receptor tyrosine phosphatase that negatively regulates an FGF receptor signaling pathway in Caenorhabditis elegans. Genes Dev. 12, 1425–1437. Abstract
Kornfeld, K., Guan, K.L., and Horvitz, H.R. (1995). The Caenorhabditis elegans gene mek-2 is required for vulval induction and encodes a protein similar to the protein kinase MEK. Genes Dev. 9, 756–768. Abstract
Kornfeld, K., Hom, D.B., and Horvitz, H.R. (1995). The ksr-1 gene encodes a novel protein kinase involved in ras-mediated signaling in Caenorhabditis elegans. Cell 83, 903–913. Abstract Article
Lackner, M.R., Kornfeld, K., Miller, L.M., Horvitz, H.R., and Kim, S.K. (1994). A MAP kinase homolog, mpk-1, is involved in ras-mediated induction of vulval cell fates in Caenorhabditis elegans. Genes Dev. 8, 160–173. Abstract
Lee, J., Jongeward, G.D., and Sternberg, P.W. (1994). unc-101, a gene required for many aspects of Caenorhabditis elegans development and behavior, encodes a clathrin-associated protein. Genes Dev. 8, 60–73. Abstract
Lu, X., and Horvitz, H.R. (1998). lin-35 and lin-53, two genes that antagonize a C. elegans Ras pathway, encode proteins similar to Rb and its binding protein RbAp48. Cell 95, 981–991. Abstract Article
Maloof, J.N., and Kenyon, C. (1998). The Hox gene lin-39 is required during C. elegans vulval induction to select the outcome of Ras signaling. Development 125, 181–190. Abstract
Malumbres, M., and Barbacid, M. (2002). RAS oncogenes: the first 30 years. Nat. Rev. Cancer 3, 7–13. Abstract Article
Miller, L.M., Gallegos, M.E., Morisseau, B.A., and Kim, S. (1993). Lin-31, a Caenorhabditis elegans HNF-3/forkhead transcription factor homolog, specificies three alternative cell fates in vulval development. Genes Dev. 7, 933–947. Abstract
Miller, M.A., Ruest, P.J., Kosinski, M., Hanks, S.K., and Greenstein, D. (2003). An Eph receptor sperm-sensing control mechanism for oocyte meiotic maturation in Caenorhabditis elegans. Genes Dev. 17, 187–200. Abstract Article
Moghal, N., Garcia, L.R., Khan, L.A., Iwasaki, K., and Sternberg, P.W. (2003). Modulation of EGF receptor-mediated vulva development by the heterotrimeric G-protein Galphaq and excitable cells in C. elegans. Development 130, 4553–4566. Abstract Article
Moghal, N., and Sternberg, P.W. (2003). The epidermal growth factor system in Caenorhabditis elegans. Exp. Cell Res. 284, 150–159. Abstract Article
Morrison, D.K., and Davis, R.J. (2003). Regulation of MAP kinase signaling modules by scaffold proteins in mammals. Annu. Rev. Cell Dev. Biol. 19, 91–118. Abstract Article
Muller, J., Ory, S., Copeland, T., Piwnica-Worms, H., and Morrison, D.K. (2001). C-TAK1 regulates Ras signaling by phosphorylating the MAPK scaffold KSR1. Mol. Cell 8, 983–993. Abstract Article
Myers, T., and Greenwald, I. (in press). lin-35 Rb acts in the major hypodermal syncitium to oppose induction of vulval precursor cells in C. elegans. Dev. Cell
Nelson, F.K., and Riddle, D.L. (1984). Functional study of the C. elegans secretory-excretory system using laser microsurgery. J. Exp. Zool. 231, 45–56. Abstract Article
Nicholas, H.R., and Hodgkin, J. (2004). The ERK MAP kinase cascade mediates tail swelling and a protective response to rectal infection in C. elegans. Curr. Biol. 14, 1256–1261. Abstract Article
Nilsson, L., Li, X., Tiensuu, T., Auty, R., Greenwald, I., and Tuck, S. (1998). Caenorhabditis elegans lin-25: cellular focus, protein expression and requirement for sur-2 during induction of vulval fates. Development 125, 4809–4819. Abstract
Nilsson, L., Tiensuu, T., and Tuck, S. (2000). Caenorhabditis elegans lin-25: A Study of Its Role in Multiple Cell Fate Specification Events Involving Ras and the Identification and Characterization of Evolutionarily Conserved Domains. Genetics 156, 1083–1096. Abstract
Ohmachi, M., Rocheleau, C.E., Church, D., Lambie, E., Schedl, T., and Sundaram, M.V. (2002). C. elegans ksr-1 and ksr-2 have both unique and redundant functions and are required for MPK-1 ERK phosphorylation. Curr. Biol. 12, 427–433. Abstract Article
Ory, S., Zhou, M., Conrads, T.P., Veenstra, T.D., and Morrison, D.K. (2003). Protein Phosphatase 2A positively regulates Ras signaling by dephosphorylating KSR1 and Raf-1 on critical 14-3-3 binding sites. Curr. Biol. 13, 1356–1364. Abstract Article
Popovici, C., Conchonaud, F., Birnbaum, D., and Roubin, R. (2004). Functional phylogeny relates LET-756 to fibroblast growth factor 9. J. Bio. Chem. 279, 40146–40152. Abstract Article
Popovici, C., Roubin, R., Coulier, F., Pontarotti, P., and Birnbaum, D. (1999). The family of Caenorhabditis elegans tyrosine kinase receptors: similarities and differences with mammalian receptors. Genome Res. 9, 1026–1039. Abstract Article
Rocheleau, C.E., Howard, R.M., Goldman, A.P., Volk, M.L., Girard, L.J., and Sundaram, M.V. (2002). A lin-45 raf enhancer screen identifies eor-1, eor-2 and unusual alleles of Ras pathway genes in Caenorhabditis elegans. Genetics 161, 121–131. Abstract
Rocheleau, C.E., Ronnlund, A., Tuck, S., and Sundaram, M.V. (2005). Caenorhabditis elegans CNK-1 promotes Raf activation but is not essential for Ras/Raf signaling. Proc. Natl. Acad. Sci. 102, 11757–11762. Abstract Article
Roubin, R., Naert, K., Popovici, C., Vatcher, G., Coulier, F., Thierry-Mieg, J., Pontarotti, P., Birnbaum, D., Baillie, D., and Thierry-Mieg, D. (1999). let-756, a C. elegans fgf essential for worm development. Oncogene 18, 6741–6747. Abstract Article
Schlessinger, J. (2000). Cell signaling by receptor tyrosine kinases [see comments]. Cell 103, 211–225. Abstract Article
Schutzman, J.L., Borland, C.Z., Newman, J.C., Robinson, M.K., Kokel, M., and Stern, M.J. (2001). The Caenorhabditis elegans EGL-15 signaling pathway implicates a DOS-like multisubstrate adaptor protein in fibroblast growth factor signal transduction. Mol. Cell Biol. 21, 8104–8116. Abstract Article
Selfors, L.M., Schutzman, J.L., Borland, C.Z., and Stern, M.J. (1998). soc-2 encodes a leucine-rich repeat protein implicated in fibroblast growth factor receptor signaling. Proc. Natl. Acad. Sci. USA 95, 6903–6908. Abstract Article
Shaye, D.D., and Greenwald, I. (2002). Endocytosis-mediated downregulation of LIN-12/Notch upon Ras activation in Caenorhabditis elegans. Nature 420, 686–690. Abstract Article
Sieburth, D.S., Sun, Q., and Han, M. (1998). SUR-8, a conserved Ras-binding protein with leucine-rich repeats, positively regulates Ras-mediated signaling in C. elegans. Cell 94, 119–130. Abstract Article
Sieburth, D.S., Sundaram, M., Howard, R.M., and Han, M. (1999). A PP2A regulatory subunit positively regulates Ras-mediated signaling during Caenorhabditis elegans vulval induction. Genes Dev. 13, 2562–2569. Abstract Article
Simske, J.S., Kaech, S.M., Harp, S.A., and Kim, S.K. (1996). LET-23 receptor localization by the cell junction protein LIN-7 during C. elegans vulval induction. Cell 85, 195–204. Abstract Article
Simske, J.S., and Kim, S.K. (1995). Sequential signalling during Caenorhabditis elegans vulval induction. Nature 375, 142–146. Abstract Article
Singh, N., and Han, M. (1995). sur-2, a novel gene, functions late in the let-60 ras-mediated signaling pathway during Caenorhabditis elegans vulval induction. Genes Dev. 9, 2251–2265. Abstract
Stern, M.J., and Horvitz, H.R. (1991). A normally attractive cell interaction is repulsive in two C. elegans mesodermal cell migration mutants. Development 113, 797–803. Abstract
Sternberg, P.W., and Han, M. (1998). Genetics of RAS signaling in C. elegans. Trends Genet. 14, 466–472. Abstract Article
Stevens, J.L., Cantin, G.T., Wang, G., Shevchenko, A., and Berk, A.J. (2002). Transcription control by E1A and MAP kinase pathway via Sur2 mediator subunit. Science 296, 755–758. Abstract Article
Sulston, J.E., and Horvitz, H.R. (1977). Post-embryonic cell lineages of the nematode Caenorhabditis elegans. Dev. Biol. 56, 110–156. Abstract Article
Sundaram, M., and Han, M. (1995). The C. elegans ksr-1 gene encodes a novel Raf-related kinase involved in Ras-mediated signal transduction. Cell 83, 889–901. Abstract Article
Sundaram, M., Yochem, J., and Han, M. (1996). A Ras-mediated signal transduction pathway is involved in the control of sex myoblast migration in Caenorhabditis elegans. Development 122, 2823–2833. Abstract
Sundaram, M.V. (2004). Vulval development: the battle between Ras and Notch. Curr. Biol. 14, R311-R313. Abstract Article
Tan, P., and Kim, S.K. (1999). Signaling specificity: the RTK/Ras/MAP kinase pathway in metazoans. Trends Genet. 15, 145–149. Abstract Article
Tan, P.B., Lackner, M.R., and Kim, S.K. (1998). MAP kinase signaling specificity mediated by the LIN-1 Ets/LIN-31 WH transcription factor complex during C. elegans vulval induction. Cell 93, 569–580. Abstract Article
Thomas, J.H., and Horvitz, H.R. (1999). The C. elegans gene lin-36 acts cell autonomously in the lin-35 Rb pathway. Development 126, 3449–3459. Abstract
Thomas, J.H., Stern, M.J., and Horvitz, H.R. (1990). Cell interactions coordinate the development of the C. elegans egg-laying system. Cell 62, 1041–1052. Abstract Article
Tuck, S., and Greenwald, I. (1995). lin-25, a gene required for vulval induction in Caenorhabditis elegans. Genes Dev. 9, 341–357. Abstract
Wang, M., and Sternberg, P. (2001). Pattern formation during C. elegans vulval induction. Curr. Top. Dev. Biol. 51, 189–220. Abstract
Wu, Y., and Han, M. (1994). Suppression of activated Let-60 Ras defines a role of Caenorhabditis elegans sur-1 MAP kinase in vulval differentiation. Genes Dev. 8, 147–159. Abstract
Wu, Y., Han, M., and Guan, K.L. (1995). MEK-2, a Caenorhabditis elegans MAP kinase kinase, functions in Ras-mediated vulval induction and other developmental events. Genes Dev. 9, 742–755. Abstract
Yin, X., Gower, N.J., Baylis, H.A., and Strange, K. (2004). Inositol 1,4,5-triphosphate signaling regulates rhythmic contractile activity of myoepithelial sheath cells in Caenorhabditis elegans. Mol. Biol. Cell 15, 3938–3949. Abstract Article
Yochem, J., Sundaram, M., and Han, M. (1997). Ras is required for a limited number of cell fates and not for general proliferation in Caenorhabditis elegans. Mol. Cell Biol. 17, 2716–2722. Abstract
Yoder, J.H., Chong, H., Guan, K.-L., and Han, M. (2004). Modulation of KSR activity in Caenorhabditis elegans by Zn ions, PAR-1 kinase and PP2A phosphatase. EMBO J. 23, 111–119. Abstract Article
Yoo, A.S., Bais, C., and Greenwald, I. (2004). Crosstalk between the EGFR and LIN-12/Notch pathways in C. elegans vulval development. Science 303, 663–666. Abstract Article
*Edited by Iva Greenwald. Last revised December 17, 2004. Published February 11, 2006. This chapter should be cited as: Sundaram, M. V. RTK/Ras/MAPK signaling (February 11, 2006), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.80.1, http://www.wormbook.org.
Copyright: © 2006 Meera V. Sundaram. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
§To whom correspondence should be addressed. E-mail: sundaram@mail.med.upenn.edu
 All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.
All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.