Embryo series courtesy of Einhard Schierenberg
Embryo series courtesy of Einhard SchierenbergTable of Contents
Abstract
Ethanol is a widely used drug whose mechanism of action, despite intensive study, remains uncertain. Biochemical and electrophysiological experiments have identified receptors and ion channels whose functions are altered at physiological concentrations of ethanol. Yet, the contribution of these potential targets to its intoxicating or behavioral effects is unclear. Unbiased forward genetic screens for resistant or hypersensitive mutants represent an attractive means of identifying the relevant molecular targets or biochemical pathways mediating the behavioral effects of neuroactive compounds. C. elegans has proven to be a particularly useful system for such studies. The behavioral effects of ethanol occur at equivalent tissue concentrations in mammals and in C. elegans, suggesting the existence of conserved drug targets in the nervous system.
This chapter reviews the results of studies directed toward determining the mechanisms of action of ethanol. Studies of the neural adaptations that occur with prolonged drug exposure are also discussed. The methods used to characterize the actions of ethanol should be applicable to the characterizations of other compounds that affect the behavior of C. elegans.
Ethanol is a widely used drug that has profound behavioral effects. Nevertheless, the fundamental mechanisms underlying these behavioral responses have remained uncertain. Similar intoxicating effects of ethanol observed in invertebrates and mammals suggest the existence of conserved targets in the nervous system.
Ethanol affects most behaviors of C. elegans. Its neurodepressive properties have provided the basis for efficient screens for resistant or hypersensitive mutants. Molecular characterization of these mutants has led to the identification of likely drug targets and biochemical pathways mediating the behavioral effects of ethanol.
C. elegans exhibits obvious behavioral changes upon acute exposure to ethanol. Exogenous ethanol produces reversible, dose-dependent effects on the body bends responsible for locomotion, speed of movement and frequency of egg laying (Davies et al., 2003). Progressive flattening of the body-bend amplitudes becomes apparent at 100–200mM ethanol and complete flattening occurs at 400–500 mM exogenous ethanol. The changes in the amplitude of the body bends correlate with a progressive decrease in the speed of movement, as determined by image analysis (see Figure 1). A similar dose-response curve is observed for egg-laying behavior when animals are maintained on agar plates. Nearly complete inhibition of both locomotion and egg laying occur at an exogenous dose of 400–500 mM ethanol. This corresponds to an internal or tissue concentration of 22–29 mM ethanol. Ethanol concentrations can readily be measured in whole animal extracts following a typical acute exposure (22 min). The concentrations are comparable to those causing intoxication in other systems. For instance, 0.1% ethanol, a common legal drinking limit, corresponds to 21.7 mM blood ethanol. Blood and brain ethanol concentrations are the same in humans. At 400–500 mM ethanol, C. elegans is still capable of movement on plates when stimulated, although the animals are uncoordinated.
|
Figure 1. Behavioral responses of C. elegans to ethanol. (A) Dose-response curve describing the inhibitory effects of ethanol on speed (•) and frequency of egg laying (□) of the wild-type (N2) strain. The measured internal concentration of ethanol in the animals for the 400 mM and 500 mM treatments was 22 ± 0.8 mM and 29 ± 0.5 mM respectively. Error bars, s.e.m. (B) Ethanol-induced decrease in body-bend amplitude during locomotion. Ethanol (200 mM) causes a small decrease in the amplitude of body bends. Complete flattening of the body bends can occur at 400- and 500-mM ethanol. The effect is most pronounced on the posterior of the body (left). Scale bar, 200 μm.
The effect of higher concentrations of ethanol on the movement of C. elegans in liquid has also been studied. (Morgan and Sedensky, 1995). Animals exhibited a steady state response to ethanol after 5 minutes of exposure. Mobility was scored for 10 sec at each concentration of ethanol after 5 minutes of exposure. EC50s were defined as the concentration in which 50% of the animals were immobile for greater than 10 sec. Under these conditions, the EC50 of N2 was 1050 +/−30 mM exogenous ethanol. Again, the immobility was reversible. This higher concentration required to inhibit movement in liquid may correspond to an anesthetic dose of ethanol in C. elegans.
Genetic screens for mutants resistant to the behavioral effects of ethanol on locomotion and egg laying led to the isolation of 29 mutants belonging to nine complementation groups (Davies et al., 2003). All of the mutants exhibiting strong resistance to the effects of ethanol on these behaviors were found to have mutations in the same gene, slo-1. slo-1 encodes a C. elegans homologue of the mammalian BK potassium channel (Wang et al., 2001). Mutations in slo-1 result in uniquely strong resistance to the effects of ethanol on both locomotion and egg laying suggesting a special relationship between BK channels and ethanol sensitivity.
The resistance of slo-1 mutants appears relatively specific. No resistance was found to other compounds that inhibit locomotion including muscimol and serotonin. No differences in the metabolism or internal concentrations of ethanol were found in slo-1 mutants, suggesting that the resistance was not due to diminished penetration of ethanol or increased ethanol metabolism. slo-1 mutations are thought to result in hyperactive neurotransmission, reflected as increased sensitivity to the acetylcholinesterase inhibitor, aldicarb. However, mutants with much stronger aldicarb hypersensitivity, such as goa-1(n363) and dgk-1(nu62), show no or comparatively little ethanol resistance. Together with the genetic results these findings suggested that the uniquely strong resistance of slo-1 was not an indirect effect.
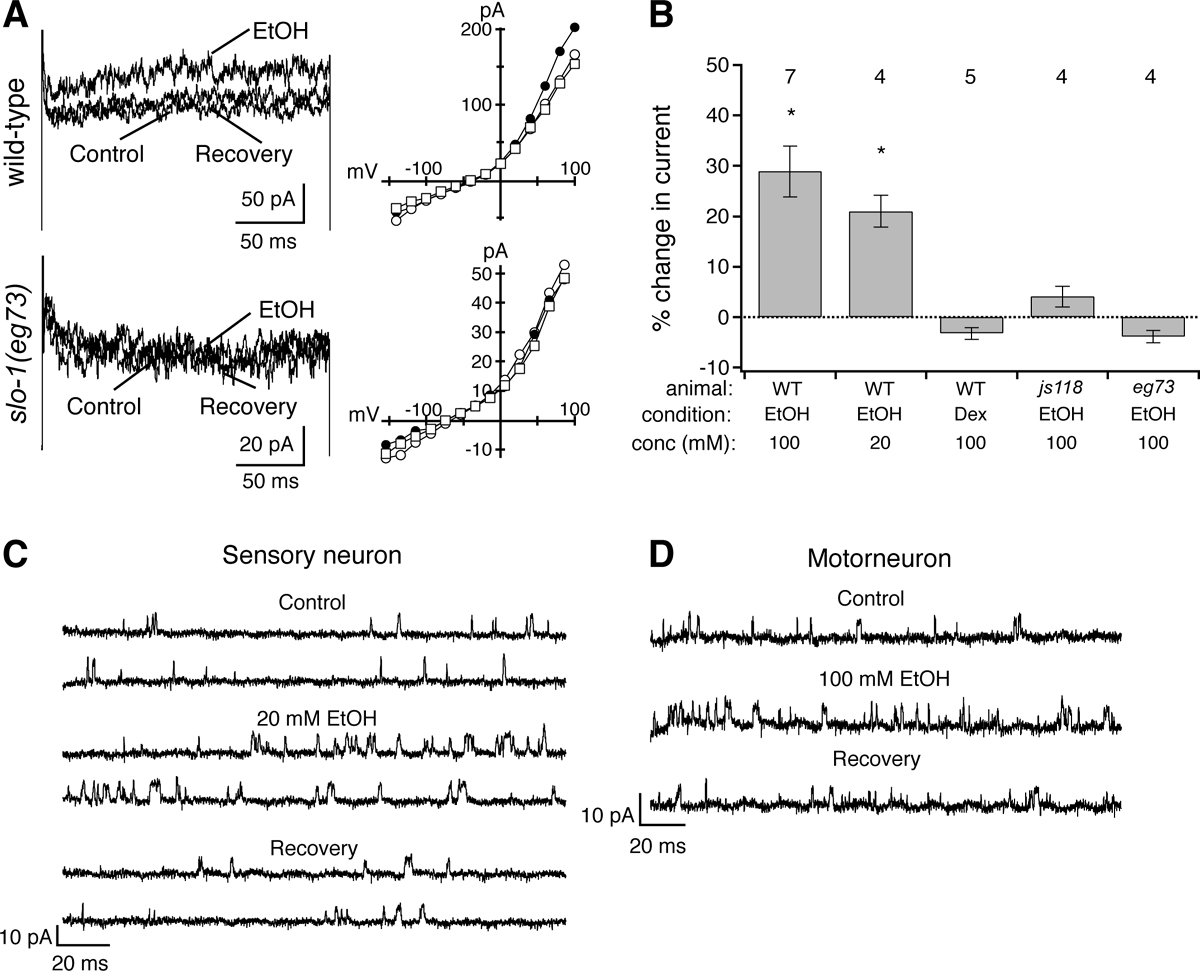
Recordings from C. elegans neurons in vivo revealed that ethanol causes an increase in a SLO-1-dependent current (see Figure 2). This effect occurs at doses of ethanol that cause intoxication in C. elegans as well as in mammalian systems. No effect of ethanol on whole-cell currents was observed in slo-1 mutants. Ethanol increased the frequency of SLO-1 channel openings in excised patches, but did not alter the size of the conductance. The ethanol effects on single channel activity occur in the absence of cytosolic factors, consistent with a direct effect of ethanol on the BK channel. Similar effects of ethanol on C. elegans SLO-1 channels can be observed in recordings from multiple different neurons including sensory neurons, interneurons and motorneurons. Activation of SLO-1 channels should inhibit neuronal excitability and decrease neurotransmitter release, possibly explaining the neurodepressive properties of ethanol.
Analysis of gain-of-function mutations in slo-1 confirmed the role of SLO-1 activation in ethanol sensitivity. Recordings from two gain-of-function mutants, ky389gf and ky399gf, revealed increases in the frequency of SLO-1 channel opening, similar in magnitude to that produced by intoxicating concentrations of ethanol. The ky389gf and ky399gf mutations result in similar depressive behavioral effects as those generated by ethanol, including comparable decreases in speed of locomotion and egg laying frequency. Hence, a selective increase in SLO-1 channel activity could recapitulate behavioral effects of ethanol. Although the role of BK channel activation in ethanol intoxication in mammals is not fully understood, similar ethanol-induced increases in the frequency of opening of mammalian BK channels have been observed in vitro (Dopico et al., 1998).
Ethanol hypersensitive mutants of C. elegans have also been described (Morgan and Sedensky, 1995). A set of mutants originally identified on the basis of hypersensitivity to volatile anesthetics was tested for altered ethanol sensitivity. At higher doses, ethanol can act as an anesthetic. Testing was completed at the higher concentrations of ethanol required to inhibit movement in liquid. Several of the mutants that were hypersensitive to volatile anesthetics (see C. elegans volatile anesthetics) also showed hypersensitivity to ethanol in this assay. One of these mutants, unc-79, was resistant rather than hypersensitive to ethanol. The mechanism of the altered responses to ethanol in this class of mutants is unclear.
|
Figure 2. Activation of the SLO-1 current by ethanol. (A) Bath application of ethanol selectively and reversibly increases the magnitude of the outward-rectifying current in wild-type
but not slo-1(-) mutant CEP neurons. Current traces for voltage steps to +86 mV holding potential on left and mean current versus voltage plots on right for before ( ), during (
), during ( ) and after (
) and after ( ) treatment with ethanol (EtOH). (B) Average percent change in whole-cell current at +86 mV is plotted for different
conditions and animals. Dextrose = Dex. Sample size indicated above each bar. Asterisks indicate significant changes (P < 0.05). (C) Ethanol reversibly potentiates the activity of a single SLO-1 channel in an excised patch from a sensory CEP neuron held at +6 mV. Dotted lines indicate baseline levels. (D) Recording
of SLO-1 channels excised from VA motorneurons also shows reversible potentiation by ethanol (100 mM) at +40 mV holding potential.
) treatment with ethanol (EtOH). (B) Average percent change in whole-cell current at +86 mV is plotted for different
conditions and animals. Dextrose = Dex. Sample size indicated above each bar. Asterisks indicate significant changes (P < 0.05). (C) Ethanol reversibly potentiates the activity of a single SLO-1 channel in an excised patch from a sensory CEP neuron held at +6 mV. Dotted lines indicate baseline levels. (D) Recording
of SLO-1 channels excised from VA motorneurons also shows reversible potentiation by ethanol (100 mM) at +40 mV holding potential.
As in other systems, acute behavioral adaptation (acute tolerance) occurs in C. elegans (Davies et al., 2004). Adaptation occurs in most systems provided that tissue concentrations of ethanol remain relatively constant. It is believed that the adaptation to ethanol represents plasticity of the nervous system or a mechanism that compensates for the physiological effects of the drug. In C. elegans, adaptation to the effect of ethanol on locomotion occurs in the absence of any change in the internal or tissue concentration of ethanol (Figure 3). Adaptation occurs more rapidly in CB4856 (a wild strain isolated in Hawaii) than N2. These differences in the rate of development of acute tolerance were found to be due to allelic variation in a single gene, npr-1. npr-1 encodes a NPY-like receptor protein (de Bono and Bargmann, 1998). Allelic variation in npr-1 had previously been shown to account for differences in food-dependent behaviors, including social (CB4856) verses solitary (N2) feeding. CB4856 has a lower function allele of npr-1 than the allele found in the N2 strain. Multiple wild isolates are known to have either the N2 or the CB4856 allele of npr-1. When these isolates were tested, the relative rate of adaptation to ethanol was predicted in all cases by the particular allele of npr-1. More rapid ethanol tolerance was also observed in animals carrying a loss of function allele of npr-1 in an otherwise N2 background. A known suppressor, ocr-2(ok47), of the food-dependent clumping behavior of npr-1 loss-of-function mutants did not suppress the more rapid adaptation to ethanol suggesting that the functions of npr-1 in ethanol tolerance and food-dependent behavior are separable. Replacing npr-1 function in different subsets of npr-1-expressing cells indicated that npr1 acts in different neurons or a larger subset of neurons for the modulation of ethanol tolerance compared with those neurons required for social behavior.
|
Figure 3. Acute ethanol tolerance in C. elegans varies between two wild strains. (A) The relative speed (% of the speed of untreated animals) of three strains increases during a continuous exposure to exogenous ethanol (500 mM). The wild strain, CB4856 and a CB4856-derived strain that had been outcrossed seven times to the N2 strain, and selected for a rapid development of acute tolerance at each outcross, demonstrate more rapid recovery of speed (acute tolerance), particularly in the 10–30 minute interval than does the N2 wild strain. Error bars, s.e.m. (B) C. elegans has low permeability to exogenous ethanol. The internal ethanol concentration (mM) within the two wild strains, N2 and CB4856, at the 10-, 30 and 50 minute time points remains constant but CB4856 demonstrated higher internal ethanol concentrations than N2.
Although it is unclear whether npr-1-related genes play a role in natural variation in ethanol responses in higher systems, there is evidence for a role of NPY signaling pathways in acute tolerance to ethanol in rodents. Knockout mutants of the NPY-encoding gene and the NPY Y1 receptor-encoding gene each results in more rapid tolerance to ethanol (Thiele et al., 1998; Thiele et al., 2002)
Kwon et al. (2004) have taken a genomics approach to understanding some of the actions of ethanol and the response of the organism. They used wild-type worms treated with an anesthetic dose of ethanol and collected RNA at various time points during the ethanol treatment. They used that RNA to probe microarrays that represent almost all of the predicted C. elegans open reading frames to determine where and when changes in gene expression occurs during ethanol treatment. They identified 230 genes that showed altered expression during the treatment, approximately 1% of the genes in C. elegans (Kwon et al., 2004). Most of these changes were found to occur by the 6 hour time point although some genes were found to respond in as little time as 15 minutes. slo-1 and npr-1 were not identified as genes that showed changes in expression; this suggests that whatever response the worm makes to compensate for the presence of ethanol in its system, it is not through direct changes in expression levels of those two genes that have been shown by mutant analysis to affect ethanol responses.
Genetic studies in C. elegans have provided a means of understanding the mechanisms of action of ethanol. Despite its profound effects on behavior, the molecular targets of ethanol have remained elusive for decades. Biochemical approaches in vitro have suggested that ethanol can affect the activity of a wide variety of neural proteins. It has been unclear whether such effects relate to the behavioral responses to these drugs.
Many of the mechanisms identified in C. elegans are likely to contribute to behavioral effects of ethanol in mammalian systems. For instance, it is intriguing that the mammalian BK channel can be modulated by ethanol in vitro in the same way that the C. elegans channel is modulated in vivo. Further genetic analysis in C. elegans may lead to a more complete understanding of both the acute and chronic effects of ethanol. It may be possible to gain a molecular understanding of not only the acute behavioral responses, but also the more complex adaptations and compensatory mechanisms that are likely to be responsible for drug tolerance, sensitization, withdrawal and dependency.
I thank Andrew Davies and Jonathan Pierce-Shimomura for their comments on the manuscript and assistance with figures.
Davies, A.G., Bettinger, J.C., Thiele, T.R., Judy, M.E., and McIntire, S.L. (2004). Natural variation in the npr-1 gene modifies ethanol responses of wild strains of C. elegans. Neuron 42, 731–743. Abstract Article
Davies, A.G., Pierce-Shimomura, J.T., Kim, H., VanHoven, M.K., Thiele, T.R., Bonci, A., Bargmann, C.I., and McIntire, S.L. (2003). A central role of the BK potassium channel in behavioral responses to ethanol in C. elegans. Cell 115, 656–666. Abstract Article
deBono, M., and Bargmann, C.I. (1998). Natural variation in a neuropeptide Y receptor homolog modifies social behavior and food response in C. elegans. Cell 94, 679–689. Abstract Article
Dopico, A.M., Anantharam, V., and Treistman, S.N. (1998). Ethanol increases the activity of Ca++ dependent K (mslo) channels: functional interaction with cystolic Ca++. J. Pharmacol. Exp. Ther. 284, 258–268. Abstract
Kwon, J.Y., Hong, M., Choi, M.S., Kang, S., Duke, K., Kim, S., Lee, S., and Lee, J. (2004). Ethanol-response genes and their regulation analyzed by a microarray and comparative genomic approach in the nematode Caenorhabditis elegans. Genomics 83, 600–614. Abstract Article
Morgan, P.G., and Sedensky, M.M. (1995). Mutations affecting sensitivity to ethanol in the nematode, Caenorhabditis elegans. Alcohol. Clin. Exp. Res. 19, 1423–1429. Abstract Article
Thiele, T.E., Koh, M.T., and Pedrazzini, T. (2002). Voluntary alcohol consumption is controlled via the neuropeptide Y Y1 receptor. J. Neurosci. 11, RC208. Abstract
*Edited by Andres V. Maricq. Last revised October 19, 2006. Published April 29, 2010. This chapter should be cited as: McIntire, S.L. Ethanol (April 29, 2010), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.40.1, http://www.wormbook.org.
Copyright: © 2010 Steven L. McIntire. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
§To whom correspondence should be addressed. E-mail: slm@gallo.ucsf.edu
 All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.
All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.