Embryo series courtesy of Einhard Schierenberg
Embryo series courtesy of Einhard SchierenbergAbstract
Wild C. elegans and other nematodes live in dirt and eat bacteria, relying on mechanoreceptor neurons (MRNs) to detect collisions with soil particles and other animals as well as forces generated by their own movement. MRNs may also help animals detect bacterial food sources. Hermaphrodites and males have 22 putative MRNs; males have an additional 46 MRNs, most, if not all of which are needed for mating. This chapter reviews key aspects of C. elegans mechanosensation, including MRN anatomy, what is known about their contributions to behavior as well as the neural circuits linking MRNs to movement. Emerging models of the mechanisms used to convert mechanical energy into electrical signals are also discussed. Prospects for future research include expanding our understanding of the molecular basis of mechanotransduction and how activation of MRNs guides and modulates behavior.
Wild C. elegans and other free-living nematodes live in dirt and eat bacteria. In this complex, three-dimensional environment, worms experience external forces produced by surface tension, by colliding with adjacent soil particles and other animals, as well as forces generated by their own movement. Hermaphrodites have 30 sensory neurons that might be recruited to detect these forces, while males have 52 additional putative mechanoreceptor neurons (MRNs; Table 1).
Table 1. Putative C. elegans mechanoreceptor neurons (MRNs).
| Hermaphrodite MRNs (30) | Male-specific MRNs (52) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| body wall | Touch receptor neurons:
|
fan | Ray Neurons:
|
||||||||||
| hook |
|
||||||||||||
| nose tip |
|
spicule |
|
||||||||||
| *SPC neurons innervate the protractor muscles in males and have been proposed to act as proprioceptors because they are attached to the protractor and anal depressor muscles (Sulston et al., 1980). | |||||||||||||
Several laboratory assays of behavioral responses to touch have been developed for C. elegans see the Behavior section in WormMethods. These assays, especially the assay for responses to gentle touch, have provided critical tools for the study of mechanosensation. As a result, more is known about the molecules needed for the function, specification, and development of C. elegans MRNs than is known in other organisms. This chapter reviews key aspects of MRN function, focusing on the anatomy of MRNs, molecules proposed to mediate the conversion of mechanical energy into electrical signals, and the neural networks linking activation of MRNs to behavior. To learn more about similarities with mechanosensation in other organisms, the reader should consult the following recent review articles: Ernstrom and Chalfie, 2002; Goodman and Schwarz, 2003; Syntichaki and Tavernarakis, 2004.
In one sense, all mechanoreceptor neurons (MRNs) are similar: they generate electrical signals in response to mechanical energy and transmit those signals to other neurons via electrical and/or chemical synapses. Yet, in another sense, they differ. Some are specialized to detect external mechanical stimuli, while others inform the nervous system about self-generated stimuli (e.g., stretch receptors). The sensory dendrites of MRNs may be ciliated or nonciliated and, if ciliated, they can be encapsulated within the cuticle or exposed to the external environment. Each neuron is located in a specific place in the body, optimized to detect forces delivered to that part of the body.
In C. elegans, touch initiates and modulates several behaviors, including locomotion (Chalfie et al., 1985; Wicks and Rankin, 1995), egg laying (Sawin, 1996), feeding (Chalfie et al., 1985; Keane and Avery, 2003), defecation (Thomas, 1990), and mating (Liu and Sternberg, 1995). The relevant MRNs are known for some, but not all of the behaviors affected by touch.
The six touch receptor neurons (ALML, ALMR, AVM, PLML, PLMR, PVM) extend long processes that innervate approximately one-half of the animal’s body length (Figure 1). Their processes are filled with unusual, 15-protofilament (15-p) microtubules that are cross-linked to one another and arrayed such that their distal ends are in close apposition to the cell membrane (Chalfie and Thomson, 1979). The 15-p microtubules are unique to the touch receptor neurons (Chalfie and Sulston, 1981) and arise from tubulins (MEC-7 and MEC-12) abundant in touch receptor neurons (Fukushige et al., 1999; Savage et al., 1989). Ultrastructural specializations are not restricted to the cytoskeleton, however. Touch receptor neurons are tightly coupled to the animal’s skin or cuticle, engulfed by hypodermal cells, and surrounded by an electron-dense extracellular matrix or ECM (Chalfie and Sulston, 1981). Because they are the only neurons whose processes are in such close contact with the cuticle along their length, touch receptor neurons are uniquely positioned to detect external forces applied to the animal’s body wall, as well as internal forces generated during locomotion.
|
Figure 1. Gross and fine morphology of the touch receptor neurons. Position of the cell bodies and neuronal processes of the touch receptor neurons, only the left side of the animal is shown (left) and electron micrograph of a PLM neuron (right). The electron micrograph is from an L4 animal, courtesy of J. Cueva, Stanford University.
Intact touch receptor neurons mediate behavioral responses to touches delivered by an eyebrow hair to the body wall. Such responses are described as simple avoidance behaviors and defined by a differential assay (see the Behavior section in WormMethods). Touch a worm anywhere along the anterior half of the body wall and it will back away (Movie 1); touch a backward moving worm along the posterior half of the body wall and it will move forward. If the animal is already moving forward, it will accelerate in response to a posterior touch. Animals that fail to respond to stimuli delivered by an eyebrow hair (gentle touch) retain sensitivity to stimuli delivered by prodding with a wire pick (harsh touch). Responses to anterior touch require ALML, AMLR, and AVM, while responses to posterior touch require PLML and PLMR (Chalfie et al., 1985; Wicks et al., 1996). A role for PVM in touch withdrawal has not been demonstrated (Chalfie et al., 1985; Wicks et al., 1996). Substrate vibrations produced by plate tap are believed to activate all touch receptor neurons simultaneously. Thus, behavioral responses to plate tap reflect a combination of the antagonistic reflexes, one initiated by anterior touch and activation of ALML/R and AVM and a second one initiated by posterior touch and activation of PLML/R. Stimulation of the anterior touch receptor neurons (ALML/R and AVM) dominates in adults— i.e. wild-type adults reverse direction in response to plate tap (Wicks et al., 1996).
Worms habituate to repeated plate tap, a process that requires intact touch receptor neurons (Wicks et al., 1996). The shorter the inter-stimulus interval, the faster habituation occurs (Rankin and Broster, 1992). Mutations that disrupt signaling by glutaminergic and dopaminergic neurons (eat-4 and cat-2, respectively) accelerate short-term habituation without affecting responses to single tap stimuli (Rankin and Wicks, 2000; Sanyal et al., 2004). These observations implicate the circuit linking touch to locomotion in the regulation of short-term habituation. The effect of cat-2 (which encodes tyrosine hydroxylase, an enzyme required for dopamine biosynthesis) is particularly intriguing and is proposed to involve neurohumoral effects of dopamine on the touch receptor neurons (Sanyal et al., 2004). Short-term behavioral habituation might also reflect de-sensitization of the touch receptor neurons. Indeed, trains of 30 stimuli are sufficient to decrease touch-evoked changes in intracellular Ca2+ (Suzuki et al., 2003). Mechanoreceptor current amplitude does not decline in response to trains of up to 60 stimuli, however (O'Hagan et al., 2005). Considered together, these results suggest that short-term behavioral habituation reflects plasticity in signaling processes subsequent to transduction in the touch receptor neurons and in the circuit linking them to locomotion.
The PVD cells have long anterior and posterior neurites that branch to form a complex dendritic network blanketing the body surface (Halevi et al., 2002; Tsalik et al., 2003). Since the PVD neurons were not completely reconstructed by White et al. (White et al., 1986), this branching pattern was not appreciated until recently. It is reminiscent of multi-dendritic sensory neurons that innervate the surface of insect larvae (Bodmer and Jan, 1987) and is consistent with the idea that PVD is a MRN. In addition to this dendritic network, PVD neurons have a ventrally directed neurite that synapses onto interneurons that control locomotion.
Strong mechanical stimuli (e.g., prodding the body wall with a wire) produce an avoidance response that persists in animals that lack touch receptor neurons. Killing the PVD neurons in such animals disrupts this behavior, suggesting that PVD neurons are high-threshold MRNs. Mutations in the mec-3 gene, which encodes a LIM domain transcription factor expressed in the touch receptor neurons and PVD, eliminate responses to both weak and strong mechanical stimuli (Way and Chalfie, 1989). mec-3 animals are insensitive to weak stimuli because their touch receptor neurons fail to differentiate properly. They may be insensitive to strong stimuli because mec-3 appears to disrupt PVD’s dendritic network (Tsalik et al., 2003). Mutations in the glr-1 ionotropic glutamate receptor also disrupt responses to strong stimuli (Hart et al., 1995), probably because loss of glr-1 affects signaling from PVD to interneurons regulating locomotion.
The extensive PVD dendritic network suggests that it could be activated by stimuli delivered anywhere along the body wall. PVD appears to detect forces greater than ~100 μN. This estimate was obtained by comparing the touch sensitivity of wild-type worms, which respond to 10 μN touches, with worms lacking function touch receptor neurons, which fail to respond to forces less than ~100 μN (Iris Chin, M.B.G., and Marty Chalfie, unpublished).
Very little is known about the molecules needed for the function of PVD, apart from the requirement for wild-type mec-3 for the formation of its extensive dendritic network. What is known is that PVD expresses mec-10 and osm-9, which are needed for cellular responses to mechanical stimuli in other C. elegans MRNs (Hilliard et al., 2005; O'Hagan et al., 2005; Suzuki et al., 2003).
The ciliated endings of the ASH neurons are located in the left and right amphids (Figure 2), where they are exposed to the external environment through a channel in the amphid sheath cell (Ward et al., 1975; White et al., 1986). ASH neurons accumulate fluorescent dyes via their exposed ciliated endings and screens for defects in dye-filling have been used to identify genes needed for cilia formation (Perkins et al., 1986).
|
Figure 2. Diagram of the position of sensilla and MRNs in the head. Abbreviations: as, amphid sensilla; il, inner labial sensillum; cs, cephalic sensillum; ol, outer labial sensilla. Adapted from Goodman and Schwarz (2003), with permission.
The ASH neurons are polymodal sensory neurons: they are needed for responses to mechanical (Kaplan and Horvitz, 1993), osmotic (Bargmann et al., 1990), and chemical stimuli (Hilliard et al., 2004; Troemel et al., 1995). All three kinds of stimuli evoke the same behavior: animals halt forward movement, back up, and then turn away, usually by executing an omega turn. ASH also contributes to avoidance of low pH (Sambongi et al., 2000) and toxic heavy metals (Hilliard et al., 2005; Sambongi et al., 1999). In addition to these acute avoidance behaviors, ASH (together with ADL) also contributes to aggregation and social feeding in npr-1 mutants (de Bono et al., 2002).
ASH was classified as a primary sensory neuron based on its morphology and identified as an MRN by laser ablation analysis. Support for this idea comes from experiments that recorded electrical responses to nose touch in AVA, an interneuron that receives direct synaptic input from ASH (Mellem et al., 2002). Additional support comes from in vivo Ca2+ imaging (Hilliard et al., 2005). Using cameleon, a genetically-encoded Ca2+ indicator (Miyawaki et al., 1997; Nagai et al., 2004), Hilliard et al. (Hilliard et al., 2005) found that nose touch produces Ca2+ transients in ASH cell bodies, but only in the presence of exogenous serotonin (5-HT). Assuming that sensitivity to nose touch is food dependent, this finding is explained by a model in which 5-HT signals the presence of food and that exogenous 5-HT is an effective substitute for food (Chao et al., 2004).
The four CEP neurons have ciliated dendrites distributed around the mouth, embedded in sensilla next to those that contain the OLQ endings (Figure 2). By contrast, the ADE and PDE neurons have ciliated dendrites embedded in the cuticle along the left and right lateral midlines. These putative MRNs are the only dopaminergic neurons in the hermaphrodite nervous system (Sulston et al., 1975).
Upon encountering a lawn of bacteria, worms slow their forward movement. This behavior requires intact CEP, ADE, and PDE neurons and dopamine synthesis (Sawin et al., 2000). As their sensory dendrites are encapsulated within the cuticle, it was proposed that CEP, ADE, and PDE sense the mechanical effects of crawling through bacteria. To test this idea, Sawin et al (Sawin et al., 2000) showed that sterile Sephadex G-200 beads elicited a slowing response that also required CEP, ADE, and PDE. Activation of CEP, ADE, and PDE could regulate locomotion by releasing dopamine, which could act extra-synaptically on touch receptor neurons (Sanyal et al., 2004) and on ventral cord motorneurons (Chase et al., 2004). The CEP and ADE neurons express trp-4, a worm homolog of Drosphila and zebrafish TRPN1 proteins needed for mechanotransduction by sensory epithelia (Sidi et al., 2003; Walker et al., 2000).
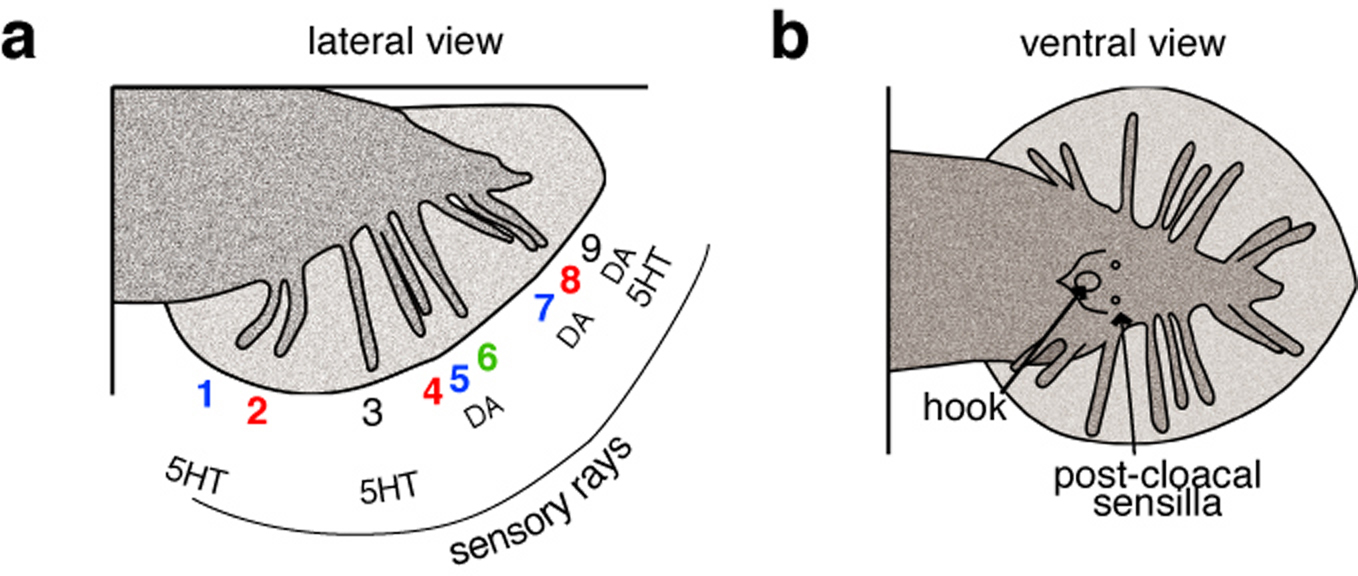
Forty-two male-specific MRNs are ciliated sensory neurons that innervate the male tail, hook, post-cloacal sensilla, and spicules (Figure 3). The hook sensilla contain two sensory neurons (HOA and HOB), while the spicule is innervated by four neurons (SPDL, SPDR, SPVL, SPVR). Thirty-six male-specific sensory neurons innervate nine bilateral pairs of sensory rays in the male tail (the RnA and RnB neurons, where ‘n’ indicates the ray number). They are exposed to the external environment in channels, except for the neurons that innervate ray 6 (which are encapsulated). Rays 1, 5, and 7 open to the dorsal side, while rays 2, 4, and 8 open ventrally. Despite looking alike, the ray sensory neurons differ with respect to their neurotransmitter phenotype. RnA in rays 5, 7, and 9 are dopaminergic, while RnB in rays 1, 3 and 9 are serotinergic. In some rays, the amine transmitters are co-expressed with the flp genes encoding FMRFamide-like peptides (Lints et al., 2004).
|
Figure 3. Sensory rays in the male tale. a. lateral view. Blue indicates rays that open to the dorsal side, red indicates rays that open to the ventral side, ray 6 is encapsulated and labeled in green. Amine neurotransmitters indicated below. b. ventral view. Adapted from Goodman and Schwarz (2003), with permission.
Male C. elegans execute a series of stereotyped behaviors during mating. Putative MRNs in the male tail play essential roles in each step of mating behavior, as determined by observing the effects of laser ablation on mating. The sensilla that innervate the ventral surface of the male tail are required for responses to ventral contact with hermaphrodites, while dorsally-directed rays mediate responses to dorsal contact with hermaphrodites (Liu and Sternberg, 1995). The hook sensilla function in vulva location, while the sensory neurons that innervate the spicule are likely to provide feedback for spicule insertion into the vulva and subsequent sperm release. It will be interesting to learn more about how the sensory information provided by the male tail is integrated and used to coordinate mating behavior.
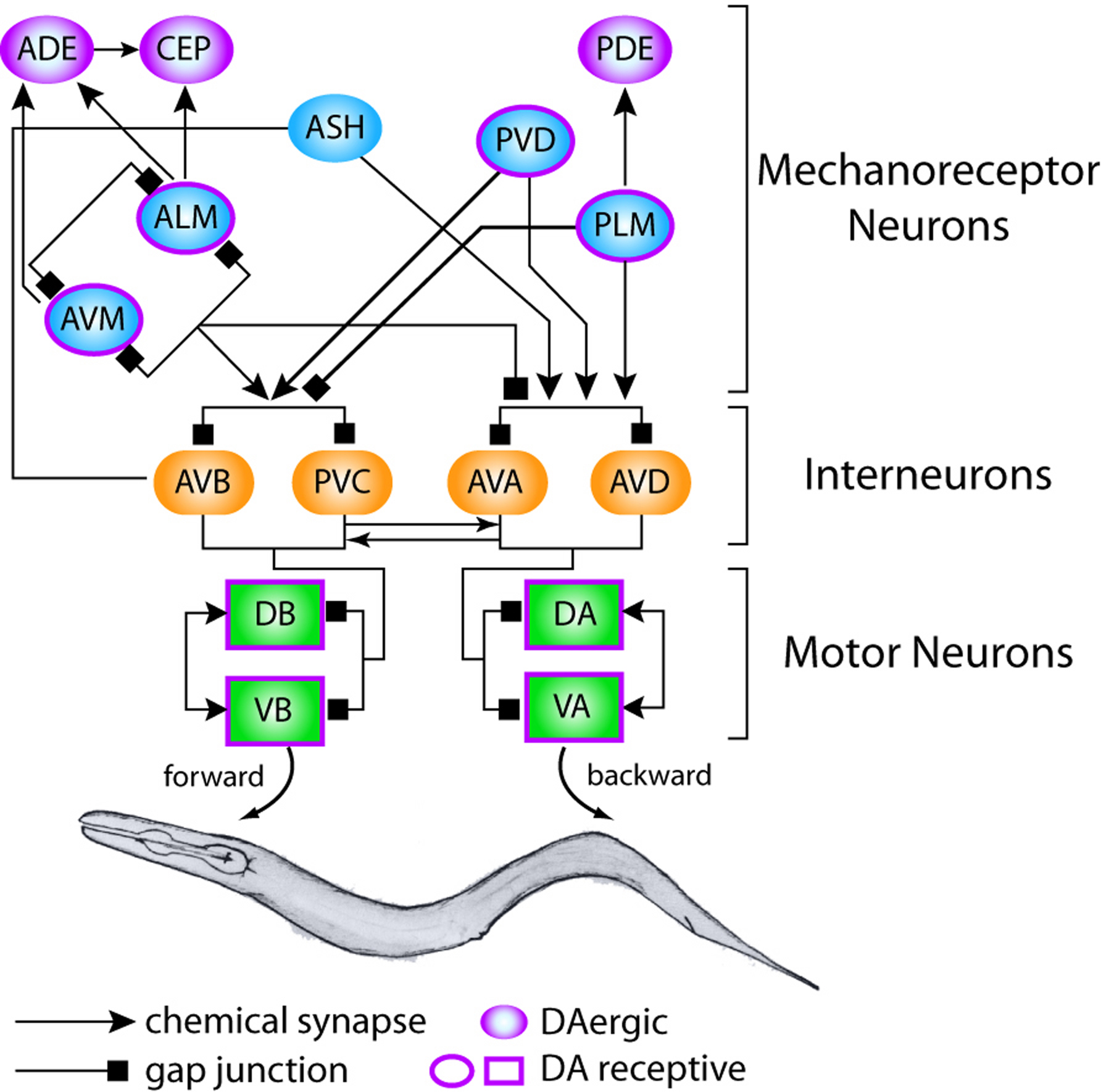
The neural circuits linking mechanosensation to locomotion in C. elegans hermaphrodites have been deduced by integrating the effects of killing individual neurons on touch-mediated behaviors (Chalfie et al., 1985; Kaplan and Horvitz, 1993; Wicks and Rankin, 1995) with the wiring diagram described in the ‘Mind of the Worm’ (White et al., 1986; Figure 4). A common thread are the four pairs of command interneurons: AVA, AVB, AVD, and PVC. AVA and AVD are needed for backward locomotion, while AVB and PVC appear to mediate forward movement.
Mechanical stimuli delivered to the nose and head elicit reversals that are frequently terminated by ‘omega turns’ (Croll, 1975). How activation of anterior MRNs leads to omega turns is unclear, however. One possibility is that MRNs signal to the SMD and RIV motor neurons, recently demonstrated to be important for omega turns (Gray et al., 2005). For stimuli that activate either the ASH neurons or the anterior touch receptor neurons, such signaling is likely to be polysynaptic, since there are no direct connections between either class of MRN and the SMD and RIV motor neurons (White et al., 1986).
|
Figure 4. Neural circuit that links mechanosensation to locomotion. MRNs are indicated by ovals, interneurons by rounded rectangles, and motor neurons by rectangles. Arrows indicate chemical synapses; squares indicate gap junctions. Dopaminergic MRNs are lavender and cells expressing predicted dopamine receptors have lavender borders.
Genetic analysis of touch sensation in worms and fruit flies has revealed subunits of putative transduction complexes (Ernstrom and Chalfie, 2002; Goodman and Schwarz, 2003). Most, if not all, proteins proposed to form transduction channels in ciliated MRNs are members of the TRP channel superfamily. TRP proteins are named for the trp (transient receptor potential) gene in Drosphila. A total of 24 C. elegans genes are predicted to encode TRP channels (Table 2: Goodman and Schwarz, 2003). This prediction has yet to be confirmed by showing that C. elegans TRPs form ion channels in heterologous cells, however. Candidate transduction channels that operate in nonciliated MRNs include members of the DEG/ENaC superfamily. Twenty-eight C. elegans genes encode members of this superfamily (Table 2; Goodman and Schwarz, 2003), named for founding members in C. elegans that can mutate to cause cellular degeneration (‘degenerins’) and vertebrate epithelial Na channels (‘ENaCs’). Three C. elegans DEG/ENaCs have been shown to form homomeric or heteromeric amiloride-sensitive Na+ channels in heterologous cells: UNC-105, MEC-4, and MEC-10 (Garcia-Anoveros et al., 1998; Goodman et al., 2002).
Table 2. Predicted C. elegans DEG/ENaC and TRP channel genes
| DEG/ENaC genes | TRP genes | |||
|---|---|---|---|---|
| CGC name | Sequence name | Subfamily | CGC name | Sequence name |
| mec-10 | F16F9.5 | TRPC | trp-1 | ZC21.2 |
| mec-4 | T01C8.7 | – | trp-2 | R06B10.4 |
| deg-1 | C47C12.6 | – | spe-41 | K01A11.4 |
| del-1 | E02H4.1 | TRPN | trp-4 | Y71A12B.4 |
| unc-8 | R13A1.4 | TRPV | osm-9 | B0212.5 |
| unc-105 | C41C4.5 | – | ocr-1 | F28H7.10 |
| flr-1 | F02D10.5 | – | ocr-2 | T09A12.3 |
| asic-1 | ZK770.1 | – | ocr-3 | T10B10.7 |
| unnamed | F26A3.6 | – | ocr-4 | Y40C5A.2 |
| unnamed | T28F2.7 | TRPA | trpa-1 | C29E6.2 |
| unnamed | F23B2.3 | – | trpa-2 | M05B5.6 |
| unnamed | C24G7.1 | TRPM | gtl-1 | C05C12.3 |
| unnamed | C24G7.2 | – | gtl-2 | F54D1.5 |
| unnamed | C24G7.4 | – | gon-2 | T01H8.5 |
| tag-324 | F28A12.1 | – | unnamed | Y38C1AB.2/6 |
| unnamed | C27C12.5 | TRPP | lov-1 | ZK945.9 |
| unnamed | T28D9.7 | – | pkd-2 | Y73F8A.1 |
| unnamed | C18B2.6 | TRPML | cup-5 | R13A5.1 |
| unnamed | C46A5.2 | elegans-specific | pqn-66 | T16A1.7 |
| unnamed | Y69H2.11 | – | ced-11 | ZK512.3 |
| unnamed | Y69H2.2 | – | unnamed | F56F11.5 |
| unnamed | Y69H2.13 | – | unnamed | W03B1.2 |
| unnamed | F55G1.12 | – | – | – |
| unnamed | T28B8.5 | – | – | – |
| asic-2 | T28F4.2 | – | – | – |
| tag-263 | C11E4.3 | – | – | – |
| unnamed | C11E4.4 | – | – | – |
| unnamed | F25D1.4 | – | – | – |
Two TRPV proteins are needed for nose touch sensation: OSM-9 and OCR-2 (Colbert et al., 1997; Tobin et al., 2002). They are co-localized in the ciliated endings of ASH, the presumed site of mechanotransduction. The proper subcellular localization of OSM-9 requires OCR-2 and vice versa, suggesting that they could form a heteromeric transduction channel. Indeed, null mutations in osm-9 abolish touch-induced Ca2+ transients in ASH (Hilliard et al., 2005). OSM-9 might also contribute to mechanotransduction in the PVD, FLP, and OLQ mechanoreceptor neurons. In OLQ, OSM-9 could form a heterologous channel with OCR-4. Nose touch avoidance is also impaired by mutations in the Δ 6 desaturase fat-3 gene. fat-3 mutants synthesize reduced amounts of several long-chain, polyunsaturated fatty acids (PUFAs), including eicosapentaenoic acid (EPA). Exogenous EPA restores nose touch sensitivity to fat-3 mutants, elicits Ca2+ transients in ASH, and can trigger avoidance responses. Both EPA-evoked Ca2+ transients and avoidance require wild-type osm-9, suggesting that EPA (and other PUFAs) act through OSM-9 channels (Kahn-Kirby et al., 2004). These findings raise the possibility that mechanical stimuli activate putative OSM-9-containing channels indirectly by mobilizing PUFAs. Additional experiments are needed to resolve this question.
Additional TRP channels that might contribute to mechanosensation include a C. elegans TRPN and TRPP protein. In Drosphila and zebrafish, TRPN1 (called NOMPC in Drosphila) is required for normal electrical responses to mechanical stimuli in mechanosensory epithelia (Sidi et al., 2003; Walker et al., 2000). trp-4 (also called CeNOMPC) encodes a TRPN protein expressed in the CEP and ADE mechanoreceptor neurons (Walker et al., 2000). No analyses of its contribution to the function of CEP and ADE in texture sensing have been reported, however. pkd-2 encodes a TRPP protein needed for male mating response and vulva location (the Lov phenotype) expressed in two classes of MRNs in the male tail (Barr et al., 2001): the ray sensory neurons and HOB neurons. PKD-2 is co-expressed with LOV-1 and, together, these proteins may form a sensory mechanotransduction channel in male-specific MRNs. The human orthologs of LOV-1 and PKD-2 appear to have this function in the primary cilium of kidney epithelial cells (Nauli et al., 2003).
The FLP, OLQ and IL1 neurons express one or more Deg/ENaC or TRP ion channel proteins, any one of which might contribute to a sensory mechanotransduction complex. FLP expresses three Deg/ENaC channel genes (mec-10, unc-8, and del-1) and one TRP channel gene (osm-9; Colbert et al., 1997; Huang and Chalfie, 1994; Tavernarakis et al., 1997). OLQ expresses two TRP channel genes (osm-9 and ocr-4), while IL1 expresses deg-1, a DEG/ENAC channel gene (Garcia-Anoveros et al., 1995; Tobin et al., 2002).
The touch receptor neurons are the best studied nonciliated MRNs in C. elegans. There is little doubt that they are primary mechanosensory neurons, since mechanical stimuli evoke increases in cytoplasmic Ca2+ as well as mechanoreceptor potentials (MRPs) and mechanoreceptor currents (MRCs; O'Hagan et al., 2005; Suzuki et al., 2003). In these cells, mechanotransduction is accomplished by activation of a DEG/ENaC ion channel complex. This MEC-4 complex is composed of four proteins that form an ion channel complex in heterologous cells (Chelur et al., 2002; Goodman et al., 2002): the degenerins MEC-4 and MEC-10, the stomatin-like protein MEC-2, and the paraoxonase-like protein MEC-6. Null mutations in mec-4, mec-2, and mec-6 eliminate both touch-evoked Ca2+ transients and MRCs (Chalfie and Au, 1989; Chalfie and Sulston, 1981; O'Hagan et al., 2005; Suzuki et al., 2003). Other ion currents expressed in touch receptor neurons are not affected by these mutations, indicating that MEC-4, MEC-2, and MEC-6 are specifically required for the generation of MRCs. Missense mutations in mec-4 and mec-10 reduce, but do not eliminate MRCs (O'Hagan et al., 2005).
Unlike ciliated MRNs, the nonciliated touch receptor neurons respond to stimuli delivered anywhere along the length of their processes. Consistent with this idea, MEC-4 (tagged with YFP) is present in puncta distributed along touch receptor neuron processes (Chelur et al., 2002). Antibodies directed against MEC-2 also stain touch receptor neuron processes in a punctate pattern. Consistent with the idea that MEC-2, MEC-4, and MEC-6 are subunits of a single ion channel complex in vivo, MEC-4::YFP puncta overlap with both MEC-2 and MEC-6::HA puncta (Chelur et al., 2002; Zhang et al., 2004). Mutations in mec-4, mec-6, or mec-10 disrupt MEC-2 puncta, suggesting that these subunits are needed to recruit MEC-2 to the complex (Zhang et al., 2004).
The distribution of MEC-4::YFP puncta is disrupted by mutations in mec-1, mec-5, mec-6 and mec-9, but not by mutations in mec-2 and mec-7 (Chelur et al., 2002; Emtage et al., 2004; Zhang et al., 2004). Thus, proper localization of the MEC-4 ion channel complex relies on some of its constituent subunits (mec-6), as well as on elements of the extracellular matrix (mec-1, mec-5, mec-9). The 15-p microtubules appear to be dispensable for both complex localization and for the generation of MRCs, since both MEC-4::YFP puncta and MRCs are present in mec-7 animals that lack 15-p microtubules (Emtage et al., 2004; O'Hagan et al., 2005). mec-1 and mec-9 encode proteins with multiple EGF and Kunitz domains, while mec-5 encodes an atypical collagen (Du et al., 1996; Emtage et al., 2004). mec-1 is also needed for attaching touch receptor processes to the cuticle (Chalfie and Sulston, 1981; Emtage et al., 2004). Mutations predicted to encode C-terminal truncated MEC-1 proteins disrupt touch sensitivity, but not process attachment (Emtage et al., 2004). Thus, touch sensitivity and process attachment are distinct processes. The observation that him-4 mutants have functional, but unattached touch receptor processes (Vogel and Hedgecock, 2001) reinforces this idea.
MEC-4, MEC-1 (fused to GFP), and MEC-5 (fused to CFP) puncta are aligned with cuticular annuli and with intermediate filaments [visualized with MH4 staining, (Francis and Waterston, 1991)] that mark hemidesmosome-like attachments in the hypodermis (Emtage et al., 2004). This observation raises the possibility that complexes that connect touch receptor neurons to the cuticle have a dual role in attachment and mechanotransduction.
Other C. elegans DEG/ENaC proteins implicated in mechanosensation include UNC-105 and UNC-8 which are expressed in body wall muscle and motor neurons, respectively (Garcia-Anoveros et al., 1998; Liu et al., 1996; Tavernarakis et al., 1997). Both channels have been proposed to act as stretch receptors—i.e. the channels are predicted to be activated by cell stretch. While initial attempts to observe such currents in wild-type muscle cells have been unsuccessful, amiloride-sensitive Na+ currents have been observed in gain-of-function unc-105 (n506) mutant muscle cells (Jospin et al., 2004). Double mutant unc-105 (n506); let-2(n281) muscle cells lack this current, consistent with the previous observation that let-2 supresses the hypercontraction phenotype of unc-10(n506) (Liu et al., 1996). No measurements of either native or expressed UNC-8-containing channels have been reported. Evidence that UNC-8 might participate in mechanosensation by motor neurons comes primarily from the observation of locomotion defects in unc-8 null mutants (Tavernarakis et al., 1997).
Much has been learned, but still more remains to be discovered about mechanosensory signaling and how such signals guide behavior. C. elegans offers an unusual opportunity to take an integrated approach to this task: existing tools permit us to study mechanosensation by observing how behavior is affected by touch, by measuring cellular responses to mechanical stimuli using in vivo Ca2+ imaging and patch-clamp recording, by searching for molecules needed for MRN function, and by manipulating cell function using cell-specific expression of engineered and heterologous proteins. Genetic screens will recover only a fraction of the genes needed for MRN function. For example, screens will miss genes with redundant or pleitropic functions. Cell-specific gene expression profiling is an alternative approach that circumvents these complications. Several new genes likely to be needed for the function of touch receptor neurons were uncovered using this approach (Zhang et al., 2002). Challenges for the future include building complete lists of the genes needed for the specification, development and function of each MRN and improved understanding of how mechanosensation affects behavior. Additional challenges involve understanding exactly how mechanical energy opens ion channels, the role of ECM components in force transfer, and delineating differences in the mechanism of mechanotransduction between ciliated and nonciliated MRNs.
I thank Erich Schwarz and two anonymous reviewers for comments; Monica Driscoll and Josh Kaplan for the sketch that inspired Figure 4. Research in my laboratory is supported by funding from the Alfred P. Sloan Foundation, the Donald B. and Delia E. Baxter Foundation, and NINDS (RO1 NS047715-01).
Bargmann, C.I., Thomas, J.H., and Horvitz, H.R. (1990). Chemosensory cell function in the behavior and development of Caenorhabditis elegans. Cold Spring Harbor Symp. Quant. Biol. 55, 529–538. Abstract
Barr, M.M., DeModena, J., Braun, D., Nguyen, C.Q., Hall, D.H., and Sternberg, P.W. (2001). The Caenorhabditis elegans autosomal dominant polycystic kidney disease gene homologs lov-1 and pkd-2 act in the same pathway. Curr. Biol. 11, 1341–1346. Abstract Article
Barr, M.M., and Sternberg, P.W. (1999). A polycystic kidney-disease gene homologue required for male mating behaviour in C. elegans. Nature 401, 386–389. Abstract Article
Bodmer, R., and Jan, Y.N. (1987). Morphological differentiation of the embryonic peripheral neurons in Drosophila. Rouxs Arch. Dev. Biol. 196, 69–77. Article
Chalfie, M., and Au, M. (1989). Genetic control of differentiation of the Caenorhabditis elegans touch receptor neurons. Science 243, 1027–1033. Abstract
Chalfie, M., and Sulston, J. (1981). Developmental genetics of the mechanosensory neurons of Caenorhabditis elegans. Dev. Biol. 82, 358–370. Abstract Article
Chalfie, M., Sulston, J.E., White, J.G., Southgate, E., Thomson, J.N., and Brenner, S. (1985). The neural circuit for touch sensitivity in Caenorhabditis elegans. J. Neurosci. 5, 956–964. Abstract
Chalfie, M., and Thomson, J.N. (1979). Organization of neuronal microtubules in the nematode Caenorhabditis elegans. J. Cell Biol. 82, 278–289. Abstract Article
Chao, M.Y., Komatsu, H., Fukuto, H.S., Dionne, H.M., and Hart, A.C. (2004). Feeding status and serotonin rapidly and reversibly modulate a Caenorhabditis elegans chemosensory circuit. Proc. Natl. Acad. Sci. USA 101, 15512–15517. Abstract Article
Chase, D.L., Pepper, J.S., and Koelle, M.R. (2004). Mechanism of extrasynaptic dopamine signaling in Caenorhabditis elegans. Nat. Neurosci. 7, 1096–1103. Abstract Article
Chelur, D.S., Ernstrom, G.G., Goodman, M.B., Yao, C.A., Chen, L.R., O.H., and Chalfie, M. (2002). The mechanosensory protein MEC-6 is a subunit of the C. elegans touch-cell degenerin channel. Nature 420, 669–673. Abstract Article
Colbert, H.A., Smith, T.L., and Bargmann, C.I. (1997). OSM-9, a novel protein with structural similarity to channels, is required for olfaction, mechanosensation, and olfactory adaptation in Caenorhabditis elegans. J. Neurosci. 17, 8259–8269. Abstract
Croll, N.A. (1975). Behavioural analysis of nematode movement. Adv. Parasitol. 13, 71–122. Abstract
de Bono, M., Tobin, D.M., Davis, M.W., Avery, L., and Bargmann, C.I. (2002). Social feeding in Caenorhabditis elegans is induced by neurons that detect aversive stimuli. Nature 419, 899–903. Abstract Article
Du, H., Gu, G., William, C.M., and Chalfie, M. (1996). Extracellular proteins needed for C. elegans mechanosensation. Neuron 16, 183–194. Abstract Article
Emtage, L., Gu, G., Hartwieg, E., and Chalfie, M. (2004). Extracellular proteins organize the mechanosensory channel complex in C. elegans touch receptor neurons. Neuron 44, 795–807. Abstract Article
Ernstrom, G.G., and Chalfie, M. (2002). Genetics of sensory mechanotransduction. Annu. Rev. Genet. 36, 411–453. Abstract Article
Francis, R., and Waterston, R.H. (1991). Muscle cell attachment in Caenorhabditis elegans. J. Cell Biol. 114, 465–479. Abstract Article
Fukushige, T., Siddiqui, Z.K., Chou, M., Culotti, J.G., Gogonea, C.B., Siddiqui, S.S., and Hamelin, M. (1999). MEC-12, an α-tubulin required for touch sensitivity in C. elegans. J. Cell Sci. 112, 395–403. Abstract
Garcia-Anoveros, J., Garcia, J.A., Liu, J.D., and Corey, D.P. (1998). The nematode degenerin UNC-105 forms ion channels that are activated by degeneration- or hypercontraction-causing mutations. Neuron 20, 1231–1241. Abstract Article
Garcia-Anoveros, J., Ma, C., and Chalfie, M. (1995). Regulation of Caenorhabditis elegans degenerin proteins by a putative extracellular domain. Curr. Biol. 5, 441–448. Abstract Article
Goodman, M.B., Ernstrom, G.G., Chelur, D.S., O'Hagan, R., Yao, C.A., and Chalfie, M. (2002). MEC-2 regulates C. elegans DEG/ENaC channels needed for mechanosensation. Nature 415, 1039–1042. Abstract Article
Goodman, M.B., and Schwarz, E.M. (2003). Transducing touch in Caenorhabditis elegans. Annu. Rev. Physiol. 65, 429–452. Abstract Article
Gray, J.M., Hill, J.J., and Bargmann, C.I. (2005). A circuit for navigation in Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA. Abstract Article
Halevi, S., McKay, J., Palfreyman, M., Yassin, L., Eshel, M., Jorgensen, E., and Treinin, M. (2002). The C. elegans ric-3 gene is required for maturation of nicotinic acetylcholine receptors. Embo J. 21, 1012–1020. Abstract Article
Hart, A.C., Sims, S., and Kaplan, J.M. (1995). Synaptic code for sensory modalities revealed by C. elegans GLR-1 glutamate receptor. Nature 378, 82–85. Abstract Article
Hilliard, M.A., Apicella, A.J., Kerr, R., Suzuki, H., Bazzicalupo, P., and Schafer, W.R. (2005). In vivo imaging of C. elegans ASH neurons: cellular response and adaptation to chemical repellents. Embo J. 24, 63–72. Abstract Article
Hilliard, M.A., Bergamasco, C., Arbucci, S., Plasterk, R.H., and Bazzicalupo, P. (2004). Worms taste bitter: ASH neurons, QUI-1, GPA-3 and ODR-3 mediate quinine avoidance in Caenorhabditis elegans. Embo J. 23, 1101–1111. Abstract Article
Huang, M., and Chalfie, M. (1994). Gene interactions affecting mechanosensory transduction in Caenorhabditis elegans. Nature 367, 467–470. Abstract Article
Jospin, M., Mariol, M.C., Segalat, L., and Allard, B. (2004). Patch clamp study of the UNC-105 degenerin and its interaction with the LET-2 collagen in Caenorhabditis elegans muscle. J. Physiol. 557, 379–388. Abstract Article
Kahn-Kirby, A.H., Dantzker, J.L., Apicella, A.J., Schafer, W.R., Browse, J., Bargmann, C.I., and Watts, J.L. (2004). Specific polyunsaturated fatty acids drive TRPV-dependent sensory signaling in vivo. Cell 119, 889–900. Abstract Article
Kaplan, J.M., and Horvitz, H.R. (1993). A dual mechanosensory and chemosensory neuron in Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 90, 2227–2231. Abstract
Keane, J., and Avery, L. (2003). Mechanosensory inputs influence Caenorhabditis elegans pharyngeal activity via ivermectin sensitivity genes. Genetics 164, 153–162. Abstract
Lints, R., Jia, L., Kim, K., Li, C., and Emmons, S.W. (2004). Axial patterning of C. elegans male sensilla identities by selector genes. Dev. Biol. 269, 137–151. Abstract Article
Liu, J., Schrank, B., and Waterston, R.H. (1996). Interaction between a putative mechanosensory membrane channel and a collagen. Science 273, 361–364. Abstract
Liu, K.S., and Sternberg, P.W. (1995). Sensory regulation of male mating behavior in Caenorhabditis elegans. Neuron 14, 79–89. Abstract Article
Mellem, J.E., Brockie, P.J., Zheng, Y., Madsen, D.M., and Maricq, A.V. (2002). Decoding of polymodal sensory stimuli by postsynaptic glutamate receptors in C. elegans. Neuron 36, 933–944. Abstract Article
Miyawaki, A., Llopis, J., Heim, R., McCaffery, J.M., Adams, J.A., Ikura, M., and Tsien, R.Y. (1997). Fluorescent indicators for Ca2+ based on green fluorescent proteins and calmodulin. Nature 388, 882–887. Abstract Article
Nagai, T., Yamada, S., Tominaga, T., Ichikawa, M., and Miyawaki, A. (2004). Expanded dynamic range of fluorescent indicators for Ca(2+) by circularly permuted yellow fluorescent proteins. Proc. Natl. Acad. Sci. USA 101, 10554–10559. Abstract Article
Nauli, S.M., Alenghat, F.J., Luo, Y., Williams, E., Vassilev, P., Li, X., Elia, A.E., Lu, W., Brown, E.M., Quinn, S.J., et al. (2003). Polycystins 1 and 2 mediate mechanosensation in the primary cilium of kidney cells. Nat Genet. Abstract Article
O'Hagan, R., Chalfie, M., and Goodman, M.B. (2005). The MEC-4 DEG/ENaC channel of Caenorhabditis elegans touch receptor neurons transduces mechanical signals. Nat. Neurosci. 8, 43–50. Abstract Article
Perkins, L.A., Hedgecock, E.M., Thomson, J.N., and Culotti, J.G. (1986). Mutant sensory cilia in the nematode C. elegans. Dev. Biol. 117, 456–487. Abstract Article
Rankin, C.H., and Broster, B.S. (1992). Factors affecting habituation and recovery from habituation in the nematode Caenorhabditis elegans. Behav. Neurosci. 106, 239–249. Abstract Article
Rankin, C.H., and Wicks, S.R. (2000). Mutations of the Caenorhabditis elegans brain-specific inorganic phosphate transporter eat-4 affect habituation of the tap-withdrawal response without affecting the response itself. J. Neurosci. 20, 4337–4344. Abstract
Sambongi, Y., Nagae, T., Liu, Y., Yoshimizu, T., Takeda, K., Wada, Y., and Futai, M. (1999). Sensing of cadmium and copper ions by externally exposed ADL, ASE, and ASH neurons elicits avoidance response in Caenorhabditis elegans. Neuroreport 10, 753–757. Abstract
Sambongi, Y., Takeda, K., Wakabayashi, T., Ueda, I., Wada, Y., and Futai, M. (2000). Caenorhabditis elegans senses protons through amphid chemosensory neurons: proton signals elicit avoidance behavior. Neuroreport 11, 2229–2232. Abstract
Sanyal, S., Wintle, R.F., Kindt, K.S., Nuttley, W.M., Arvan, R., Fitzmaurice, P., Bigras, E., Merz, D.C., Hebert, T.E., van der Kooy, D., et al. (2004). Dopamine modulates the plasticity of mechanosensory responses in Caenorhabditis elegans. Embo J. 23, 473–482. Abstract Article
Savage, C., Hamelin, M., Culotti, J.G., Coulson, A., Albertson, D.G., and Chalfie, M. (1989). mec-7 is a β-tubulin gene required for the production of15-protofilament microtubules in Caenorhabditis elegans. Genes & Development 3, 870–887. Abstract
Sawin, E.R. (1996). Genetic and cellular analysis of modulated behaviors in Caenorhabditis elegans (Cambridge, MA: Massachusetts Institute of Technology)
Sawin, E.R., Ranganathan, R., and Horvitz, H.R. (2000). C. elegans locomotory rate is modulated by the environment through a dopaminergic pathway and by experience through a serotonergic pathway. Neuron 26, 619–631. Abstract Article
Sidi, S., Friedrich, R.W., and Nicolson, T. (2003). NompC TRP channel required for vertebrate sensory hair cell mechanotransduction. Science 301, 96–99. Abstract Article
Sulston, J., Dew, M., and Brenner, S. (1975). Dopaminergic neurons in the nematode Caenorhabditis elegans. J. Comp. Neurol. 163, 215–226. Abstract Article
Sulston, J.E., Albertson, D.G., and Thomson, J.N. (1980). The Caenorhabditis elegans male: postembryonic development of nongonadal structures. Dev. Biol. 78, 542–576. Abstract Article
Suzuki, H., Kerr, R., Bianchi, L., Frokjaer-Jensen, C., Slone, D., Xue, J., Gerstbrein, B., Driscoll, M., and Schafer, W.R. (2003). In vivo imaging of C. elegans mechanosensory neurons demonstrates a specific role for the MEC-4 channel in the process of gentle touch sensation. Neuron 39, 1005–1017. Abstract Article
Syntichaki, P., and Tavernarakis, N. (2004). Genetic models of mechanotransduction: the nematode Caenorhabditis elegans. Physiol. Rev. 84, 1097–1153. Abstract Article
Tavernarakis, N., Shreffler, W., Wang, S., and Driscoll, M. (1997). unc-8, a DEG/ENaC family member, encodes a subunit of a candidate mechanically gated channel that modulates C. elegans locomotion. Neuron 18, 107–119. Abstract Article
Thomas, J.H. (1990). Genetic analysis of defecation in Caenorhabditis elegans. Genetics 124, 855–872. Abstract
Tobin, D., Madsen, D., Kahn-Kirby, A., Peckol, E., Moulder, G., Barstead, R., Maricq, A., and Bargmann, C. (2002). Combinatorial expression of TRPV channel proteins defines their sensory functions and subcellular localization in C. elegans neurons. Neuron 35, 307–318. Abstract Article
Troemel, E.R., Chou, J.H., Dwyer, N.D., Colbert, H.A., and Bargmann, C.I. (1995). Divergent seven transmembrane receptors are candidate chemosensory receptors in C. elegans. Cell 83, 207–218. Abstract Article
Tsalik, E.L., Niacaris, T., Wenick, A.S., Pau, K., Avery, L., and Hobert, O. (2003). LIM homeobox gene-dependent expression of biogenic amine receptors in restricted regions of the C. elegans nervous system. Dev. Biol. 263, 81–102. Abstract Article
Vogel, B.E., and Hedgecock, E.M. (2001). Hemicentin, a conserved extracellular member of the immunoglobulin superfamily, organizes epithelial and other cell attachments into oriented line-shaped junctions. Development 128, 883–894. Abstract
Walker, R.G., Willingham, A.T., and Zuker, C.S. (2000). A Drosophila mechanosensory transduction channel. Science 287, 2229–2234. Abstract Article
Ward, S., Thomson, N., White, J.G., and Brenner, S. (1975). Electron microscopical reconstruction of the anterior sensory anatomy of the nematode Caenorhabditis elegans. J. Comp. Neurol. 160, 313–337. Abstract Article
Way, J.C., and Chalfie, M. (1989). The mec-3 gene of Caenorhabditis elegans requires its own product for maintained expression and is expressed in three neuronal cell types. Genes Dev. 3, 1823–1833. Abstract
White, J.G., Southgate, E., Thomson, J.N., and Brenner, S. (1986). The structure of the nervous system of the nematode C. elegans. Phil. Trans. Roy. Soc. B 314, 1–340. Abstract
Wicks, S.R., and Rankin, C.H. (1995). Integration of mechanosensory stimuli in Caenorhabditis elegans. J. Neurosci. 15, 2434–2344. Abstract
Wicks, S.R., Roehrig, C.J., and Rankin, C.H. (1996). A dynamic network simulation of the nematode tap withdrawal circuit: predictions concerning synaptic function using behavioral criteria. J. Neurosci. 16, 4017–4031. Abstract
*Edited by Erik M. Jorgensen and Joshua M. Kaplan. Last revised April 23, 2005. Published January 06, 2006. This chapter should be cited as: Goodman, M.B. Mechanosensation (January 06, 2006), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.62.1, http://www.wormbook.org.
Copyright: © 2006 Miriam B. Goodman. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
§To whom correspondence should be addressed. E-mail: mbgoodmn@stanford.edu
 All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.
All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.