Embryo series courtesy of Einhard Schierenberg
Embryo series courtesy of Einhard SchierenbergAbstract
Form follows function, and this maxim is particularly true for the nematode sperm cell. Motility is essential for fertilization, and the process of spermatogenesis culminates in the production of a crawling spermatozoon with an extended pseudopod. However, the morphological similarity to amoeboid cells of other organisms is not conserved at the molecular level. Instead of utilizing the actin cytoskeleton and motor proteins, the pseudopod moves via the regulated assembly and disassembly of filaments composed of the major sperm protein (MSP). The current work reviews the structure and dynamics of MSP filament formation, the critical role of pH in MSP assembly, and the recent identification of components that regulate this process. The combination of cytological, biochemical, and genetic approaches in this relatively simple system make nematode sperm an attractive model for investigating the mechanics of amoeboid cell motility.
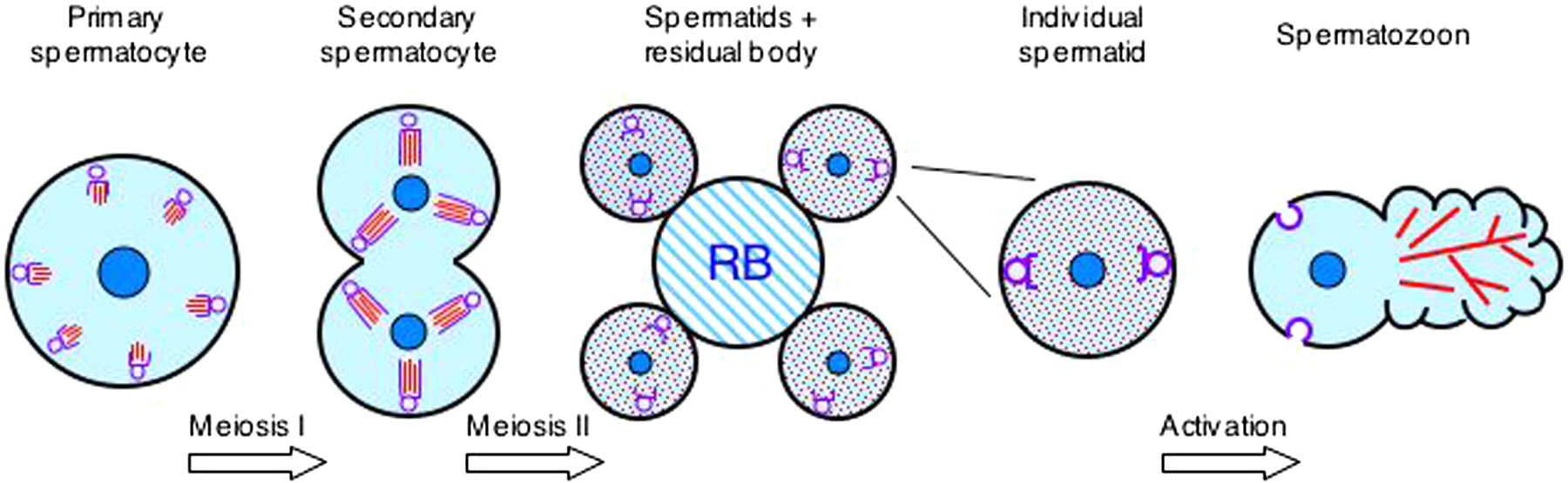
A detailed description of sperm development, or spermatogenesis, can be found in the WormBook chapter by Steve L'Hernault (see Spermatogenesis); however, a brief summary is included here and diagrammed in Figure 1 to emphasize the particulars of MSP assembly and segregation. Spermatogonial stem cells give rise to primary spermatocytes (Ward et al., 1981). The first meiotic division produces two secondary spermatocytes and the second meiosis yields four haploid spermatids. These round, non-motile spermatids bud from the surface of the residual body, which contains components that are not needed for further development or sperm function. An extracellular signal induces the process of activation, or spermiogenesis, and causes pseudopod extension and motility that is the hallmark of mature spermatozoa (Figure 2).
|
Figure 1. Spermatogenesis and MSP. Shown are various stages in sperm cell development. In primary spermatocytes, MSP (red) begins to assemble as paracrystalline arrays in fibrous bodies associated with the membranous organelles (purple). Assembly continues through the secondary spermatocyte stage. The fibrous body-membranous organelle complexes segregate into the spermatids and, following separation from the residual body (RB), MSP dissociates into the cytosol. Upon activation, MSP reassembles into filamentous fibers in the pseudopod.
|
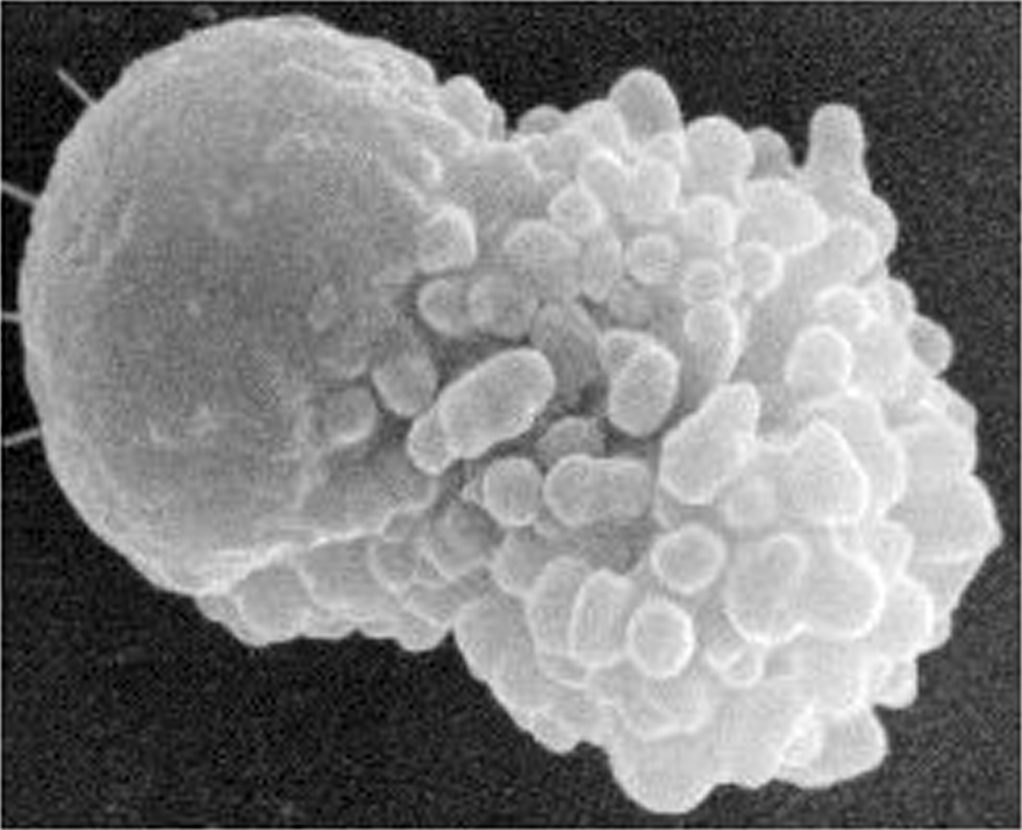
Figure 2. Scanning electron micrograph of spermatozoon. The cell body is to the left, and the numerous finger-like projections of the pseudopod to the right.
MSP localization reflects the various stages of spermatogenesis. Monoclonal antibodies to MSP first detect the protein in primary spermatocytes within structures termed membranous organelles. MSP accumulates in a paracrystalline arrays of filaments, called fibrous bodies, to form the associated fibrous body-membranous organelle complexes. These complexes segregate into the budding spermatids. Shortly after the spermatids separate from the residual body, the fibrous bodies dissociate and MSP is distributed throughout the cytoplasm (Roberts et al., 1986). After activation, MSP becomes relocalized to the pseudopod. Ultrastructural analysis (described below in greater detail) shows a complex network of MSP filaments and fibers within the pseudopod.
The mechanics of fertilization in the nematode reproductive tract demonstrate the importance of sperm motility in this process. Crawling spermatozoa accumulate in the sperm storage organ known as the spermatheca, which is the site of fertilization. As each oocyte passes through the spermatheca, many spermatozoa are dislodged into the uterus but quickly return to the spermatheca. The reproductive period spans several days and ~300 progeny are produced during this time. Essentially every spermatozoon is successful at fertilizing an oocyte, so sperm motility must be maintained throughout this interval (Ward and Carrel, 1979).
Mutational screens for sperm-specific sterility, the Fer or Spe phenotype, have identified a large number of loci necessary for proper sperm function (e.g., see Argon and Ward, 1980; L'Hernault et al., 1988). Mutations in several of the Spe/Fer genes produce apparently normal spermatids that fail to undergo spermiogenesis. These immotile cells initially localize to the spermatheca. However, the passage of oocytes rapidly sweeps the mutant sperm into the uterus, where they are unable to return to the spermatheca. Motility is not sufficient to insure functionality, since mutations in several other Spe/Fer genes permit normal activation but not fertilization; however, motility is likely a necessary prerequisite.
The Spe-8 class of mutations is particularly instructive because sex-specific differences in the activation pathway permit the in vivo manipulation of motility. Mutations of the Spe-8 class, consisting of spe-8, spe-12, spe-27, and spe-29, block activation of spermatids in hermaphrodites but not males (L'Hernault et al., 1988; Minniti et al., 1996; Nance et al., 2000). As a consequence, hermaphrodites are self-sterile while males remain cross-fertile. However, Spe-8 class hermaphrodite spermatids can be activated by mating, through transfer of a male component of the activation signal. These trans-activated sperm become motile and capable of fertilization, indicating the importance of motility for sperm cell function.
Video microscopy of crawling sperm (Movie 1) reveals the critical role of MSP polymerization in cell motility (Roberts and Ward, 1982; Sepsenwol et al., 1989). Much of this work is on the large sperm from the parasitic nematode Ascaris suum, but the same features hold true for C. elegans. Movement occurs by extension of the leading edge of the pseudopod, attachment to the substrate, and retraction of the cell body. A meshwork of fibers extends throughout the pseudopod. Assembly of new fibers at the leading edge of the pseudopod and disassembly at its base produce a treadmilling motion, and the rate of treadmilling correlates precisely with the rate of crawling (Roberts and King, 1991). Manipulation of the rates of assembly and disassembly by pH (see following section) clearly demonstrates that the motive force is coupled to the combination of these two opposing processes (Italiano et al., 1999). In other types of amoeboid cells, motility is based on the actin cytoskeleton. However, nematode spermatozoa contain no actin (Nelson et al., 1982), but instead employ MSP filaments in this role.
In vitro activation studies reveal a critical role for pH in regulating spermiogenesis. Agents that increase the intracellular pH (e.g., the weak base TEA or the ionophore monensin at basic pH) induce spermatid activation (Ward et al., 1983). The molecular mechanism of this activation is unknown; however, it is likely physiologically relevant, since in vitro activation by TEA produces mature spermatozoa capable of oocyte fertilization upon artificial insemination (LaMunyon and Ward, 1994). Indeed, initiation of the spermiogenesis pathway might culminate in elevated pH, since activation by TEA or monensin bypasses the requirement for the Spe-8 class signaling components (Shakes and Ward, 1989).
Changes in intracellular pH appear to control not just activation but also MSP polymerization throughout spermatogenesis (King et al., 1994). Experiments in A. suum show that intracellular pH is highest in spermatocytes, when MSP is assembled into paracrystalline arrays in fibrous bodies, and lowest in spermatids, when MSP disassembles into the cytosol. Spermiogenesis induces an increase in pH and the assembly of MSP macrofibers. Remarkably, a pH gradient forms within the pseudopod of the spermatozoon; a higher pH is observed at the leading edge, where new filaments assemble, than at the base, where disassembly occurs. Decreasing the pH by buffer treatment can either slow or halt MSP polymerization at the leading edge of the pseudopod without affecting disassembly. A return to physiological buffer rapidly restores MSP assembly, pseudopod extension, and motility (Italiano et al., 1999). Thus, MSP assembly in all case s is accompanied by an increase in intracellular pH. In vitro filament assembly occurs across a wide range of pH, so direct regulation of MSP polymerization by pH is unlikely (King et al., 1992); rather, pH probably alters the activity of membrane components that control the assembly and disassembly of MSP filaments.
MSP polymerization occurs through a series of increasingly higher-order interactions (King et al., 1994). Pairs of MSP monomers produce dimers, dimers assemble into helical subfilaments, pairs of subfilaments entwine to form helical filaments, and filaments associate into larger structures called fibers, macrofibers, or bundles. MSP dimers are extremely stable in solution (Kd < 5 × 10-8) and exhibit the same structure as in the filamentous form (Haaf et al., 1996); therefore, the polymerization of MSP is likely regulated at the level of subfilament assembly.
Structural studies have provided insights into the mechanics of MSP assembly that generate the motive force. X-ray diffractions of MSP crystals from both A. suum (Bullock et al., 1996) and C. elegans (Baker et al., 2002) reveal an immunoglobulin-like fold comprised of a seven-stranded β sandwich (Figure 4). MSP forms symmetrical dimers through β strands a2 and b plus the penultimate asparagine residue. The assembly of dimers into subfilaments occurs through hydrogen bonding between residues within strand g. Filament formation between subfilaments is mediated through multiple sites of interaction, and the same sites might function in filament bundling as well. Mutational and biochemical analyses of recombinant MSP have confirmed the importance of residues critical for both the immunoglobulin fold and the dimerization interface (Smith and Ward, 1998).
|
Figure 4. Structure of C. elegans MSP. (A) MSP monomer. Backbone trace demonstrates the seven-stranded immunoglobulin-like fold. (B) MSP dimer. Sidechains involved in dimer formation are shown in red. (C) MSP subfilament. Sidechains involved in subfilament interaction between two dimers are shown in green. Structure coordinates are available as entry 1GRW from the Protein Data Bank.
Purified MSP exists as a dimer under most buffer conditions; however, polymerization occurs in the presence of water-miscible alcohols (King et al., 1992). Transmission electron micrographs of these negatively stained filaments are indistinguishable from those obtained from fixed spermatozoa, suggesting that filament formation is an intrinsic property of MSP even in the absence of additional proteins.
A crucial advance in our understanding of MSP-based motility has been the development of a reconstituted cell-free filament assembly system (Italiano et al., 1996). Addition of ATP to lysate from A. suum sperm causes the assembly of MSP into fibers. Fractionation of the lysate demonstrates that both membrane and cytosolic components, in addition to MSP and ATP, are necessary for fiber formation. Phase-contrast video microscopy reveals that each MSP fiber is nucleated by a membrane vesicle, and that the vesicle is propelled forward by the growing filament. Furthermore, only the subset of vesicles that are derived from the leading edge of the pseudopod, the site of MSP polymerization in vivo, can promote MSP filament assembly in vitro. The maximum observed rate of fiber growth is similar to the rate of sperm crawling, suggesting that all of the relevant components are present.
The disassembly of MSP fibers can be promoted in the same cell-free system (Miao et al., 2003). Following fiber growth from reconstituted components, removal of ATP and addition of a tyrosine phosphatase causes fiber retraction. A bead attached to the non-vesicle end of the MSP fiber is drawn toward the vesicle end as the fiber shortens. The phosphatase likely controls the activity of a disassembly factor, since MSP itself is not phosphorylated. Thus, the reconstituted system exhibits the primary hallmarks of in vivo motility: the assembly of MSP fibers at the leading edge of the pseudopod membrane generates a motive force that propels the membrane forward, and disassembly causes fiber retraction at the trailing edge.
Investigators have used the reconstituted system to identify specific proteins that regulate the assembly of MSP. One critical component from the vesicle fraction is a 48kDa integral membrane protein with the unwieldy name major sperm protein polymerization organizing protein or, more simply, MPOP (LeClaire et al., 2003). Phosphorylation of tyrosine residues appears to regulate the activity of MPOP. Immunolabeling of spermatozoa with anti-MPOP antibodies indicates that the protein is evenly distributed throughout the sperm plasma membrane, but anti-phosphotyrosine labeling of the membrane is restricted to the leading edge of the pseudopod. Purified phosphorylated MPOP, when combined with cytosol and ATP, is sufficient to promote MSP filament assembly in the absence of other vesicle components. Intriguingly, lowering the pH blocks both tyrosine phosphorylation of MPOP and the in vivo assembly of MSP in a similar manner. This obs ervation suggests an appealing mechanism for the regulation of motility by pH, whereby an as-yet-unidentified, pH-sensitive tyrosine kinase phosphorylates MPOP at the tip of the pseudopod to nucleate the polymerization of MSP fibers.
Additional proteins have been isolated from the cytosolic fraction as well. MFP1 (for MSP fiber protein) is a complex of three related, 15-16 kDa proteins (α, β, and γ) that inhibits fiber growth (Buttery et al., 2003). Homologs in C. elegans comprise a larger gene family called SSP, for sperm-specific protein. The crystal structure of one SSP, encoded by ssp-19, is remarkably similar to MSP despite having less than 20% amino acid sequence identity (Schormann et al., 2004).
MFP2 is a 38 kDa protein that appears to be essential for fiber formation (Buttery et al., 2003). Anti-MFP2 antibodies abrogate filament assembly in the reconstituted system, and addition of purified MFP2 can stimulate assembly. Both anti-MFP1 and anti-MFP2 antibodies label MSP fibers in vivo and in vitro, suggesting that both proteins play fairly direct roles in regulating assembly. Neither protein can substitute for the cytosolic fraction in the reconstituted system, so additional components remain to be identified.
MSP-based sperm motility appears to be conserved among nematodes. Crawling spermatozoa are rare among metazoans, but they are a characteristic feature of this phylum. In C. elegans, MSP is encoded by a family of 28 genes (not including several additional pseudogenes) that produce isoforms that are 97-100% identical (Burke and Ward, 1983; Klass et al., 1984; Ward et al., 1988). MSP genes have been identified across widely diverged nematode species (e.g., see Scott et al., 1989) and MSP gene expression is restricted to the sperm (Klass et al., 1982; Ward, 1987). MSPs from all nematodes are greater than 60% identical, with particularly high sequence conservation in the dimerization interface as well as β strands f and g (the proposed site of subfilament assembly). Missense mutations that abolish MSP-MSP interaction in the yeast two-hybrid system map almost exclusively to these regions, suggesting functional conservation as well (Smith and Ward, 1998). However, mutant MSPs have not yet been characterized in the reconstituted filament assembly system, and conservation might reflect the roles of MSP in oocyte maturation or ovulation signaling rather than motility (see Control of oocyte meiotic maturation and fertilization).
MSP seems to be restricted to nematodes, so how did this unique mechanism of cell motility arise? Although homologs have not been found in other phyla, proteins with limited sequence similarity have been identified in species from plants to mammals. The first of these, VAP-33, was identified from Aplysia californica as a VAMP/synaptobrevin-interacting protein involved in neurotransmitter release from synaptic vesicles (Skehel et al., 1995). Subsequent studies in other organisms indicate a more general role in vesicle fusion and/or trafficking, and find the protein associated with microtubules and the endoplasmic reticulum in addition to vesicles (Skehel et al., 2000). All of the VAP-33 homologs are predicted to share a similar architecture: an amino-terminal MSP-like domain, a central coiled-coil region, and a carboxy-terminal transmembrane domain (Laurent et al., 2000). In addition to MSP, the C. elegans genome encodes several of these VAP-33 homologs. Thus, it appears that the small, cytosolic MSP arose from a larger, membrane-tethered vesicle protein.
Despite the apparent novelty of MSP and sperm motility, the system can offer insights into the canonical, actin-based mechanism of cell movement. The generation of force and membrane protrusion by protein polymerization is a common feature of both systems. Actin is required for multiple cellular processes, which greatly complicates the analysis of its role in motility. In contrast, the relative simplicity of sperm motility offers an attractive model for investigation, and the combination of genetic and biochemical approaches will continue to advance our understanding of the underlying biomechanical properties of amoeboid movement.
Argon, Y., and Ward, S. (1980). Caenorhabditis elegans fertilization-defective mutants with abnormal sperm. Genetics 96, 413–433. Abstract
Baker, A.M., Roberts, T.M., and Stewart, M. (2002). 2.6 Å resolution crystal structure of helices of the motile major sperm protein (MSP) of Caenorhabditis elegans. J. Mol. Biol. 319, 491–499. Abstract Article
Bullock, T.L., Roberts, T.M., and Stewart, M. (1996). 2.5 Å resolution crystal structure of the motile major sperm protein (MSP) of Ascaris suum. J. Mol. Biol. 263, 284–296. Abstract Article
Burke, D.J., and Ward, S. (1983). Identification of a large multigene family encoding the major sperm protein of Caenorhabditis elegans. J. Mol. Biol. 171, 1–29. Abstract
Buttery, S.M., Ekman, G.C., Seavy, M., Stewart, M., and Roberts, T.M. (2003). Dissection of the Ascaris sperm motility machinery identifies key proteins involved in major sperm protein-based amoeboid locomotion. Mol. Biol. Cell 14, 5082–5088. Abstract Article
Haaf, A., Butler, P.J., Kent, H.M., Fearnley, I.M., Roberts, T.M., Neuhaus, D., and Stewart, M. (1996). The motile major sperm protein (MSP) from Ascaris suum is a symmetric dimer in solution. J. Mol. Biol. 260, 251–260. Abstract Article
Italiano, J.E., Jr., Roberts, T.M., Stewart, M., and Fontana, C.A. (1996). Reconstitution in vitro of the motile apparatus from the amoeboid sperm of Ascaris shows that filament assembly and bundling move membranes. Cell 84, 105–114. Abstract Article
Italiano, J.E., Jr., Stewart, M., and Roberts, T.M. (1999). Localized depolymerization of the major sperm protein cytoskeleton correlates with the forward movement of the cell body in the amoeboid movement of nematode sperm. J. Cell Biol. 146, 1087–1096. Abstract Article
King, K.L., Essig, J., Roberts, T.M., and Moerland, T.S. (1994). Regulation of the Ascaris major sperm protein (MSP) cytoskeleton by intracellular pH. Cell Motil. Cytoskeleton 27, 193–205. Abstract Article
King, K.L., Stewart, M., and Roberts, T.M. (1994). Supramolecular assemblies of the Ascaris suum major sperm protein (MSP) associated with amoeboid cell motility. J. Cell Sci. 107, 2141–2149. Abstract Article
King, K.L., Stewart, M., Roberts, T.M., and Seavy, M. (1992). Structure and macromolecular assembly of two isoforms of the major sperm protein (MSP) from the amoeboid sperm of the nematode, Ascaris suum. J. Cell Sci. 101, 847–857. Abstract
Klass, M.R., Dow, B., and Herndon, M. (1982). Cell-specific transcriptional regulation of the major sperm protein in Caenorhabditis elegans. Dev. Biol. 93, 152–164. Abstract Article
Klass, M.R., Kinsley, S., and Lopez, L.C. (1984). Isolation and characterization of a sperm-specific gene family in the nematode Caenorhabditis elegans. Mol. Cell. Biol. 4, 529–537. Abstract
LaMunyon, C.W., and Ward, S. (1994). Assessing the viability of mutant and manipulated sperm by artificial insemination of Caenorhabditis elegans. Genetics 138, 689–692. Abstract
Laurent, F., Labesse, G., and de Wit, P. (2000). Molecular cloning and partial characterization of a plant VAP33 homologue with a major sperm protein domain. Biochem. Biophys. Res. Commun. 270, 286–292. Abstract Article
LeClaire, L.L. III, Stewart, M., and Roberts, T.M. (2003). A 48 kDa integral membrane phosphoprotein orchestrates the cytoskeletal dynamics that generate amoeboid cell motility in Ascaris sperm. J. Cell Sci. 116, 2655–2663. Abstract Article
L'Hernault, S.W., Shakes, D.C., and Ward, S. (1988). Developmental genetics of chromosome I spermatogenesis- defective mutants in the nematode Caenorhabditis elegans. Genetics 120, 435–452. Abstract
Miao, L., Vanderlinde, O., Stewart, M., and Roberts, T.M. (2003). Retraction in amoeboid cell motility powered by cytoskeletal dynamics. Science 302, 1405–1407. Abstract Article
Minniti, A.N., Sadler, C., and Ward, S. (1996). Genetic and molecular analysis of spe-27, a gene required for spermiogenesis in Caenorhabditis elegans hermaphrodites. Genetics 143, 213–223. Abstract
Nance, J., Davis, E.B., and Ward, S. (2000). spe-29 encodes a small predicted membrane protein required for the initiation of sperm activation in Caenorhabditis elegans. Genetics 156, 1623–1633. Abstract
Nelson, G.A., Roberts, T.M., and Ward, S. (1982). Caenorhabditis elegans spermatozoan locomotion: amoeboid movement with almost no actin. J. Cell Biol. 92, 121–131. Abstract Article
Roberts, T.M., and King, K.L. (1991). Centripetal flow and directed reassembly of the major sperm protein (MSP) cytoskeleton in the amoeboid sperm of the nematode, Ascaris suum. Cell Motil. Cytoskeleton 20, 228–241. Abstract Article
Roberts, T.M., Pavalko, F.M., and Ward, S. (1986). Membrane and cytoplasmic proteins are transported in the same organelle complex during nematode spermatogenesis. J. Cell Biol. 102, 1787–1796. Abstract Article
Roberts, T.M., and Ward, S. (1982). Centripetal flow of pseudopodial surface components could propel the amoeboid movement of Caenorhabditis elegans spermatozoa. J. Cell Biol. 92, 132–138. Abstract Article
Schormann, N., Symersky, J., and Luo, M. (2004). Structure of sperm-specific protein SSP-19 from Caenorhabditis elegans. Acta Crystallogr. D Biol. Crystallogr. 60, 1840–1845. Abstract Article
Scott, A.L., Dinman, J., Sussman, D.J., and Ward, S. (1989). Major sperm protein and actin genes in free-living and parasitic nematodes. Parisitology 98, 471–478. Abstract
Sepsenwol, S., Ris, H., and Roberts, T.M. (1989). A unique cytoskeleton associated with crawling in the amoeboid sperm of the nematode, Ascaris suum. J. Cell Biol. 108, 55–66. Abstract Article
Shakes, D.C., and Ward, S. (1989). Initiation of spermiogenesis in C. elegans: a pharmacological and genetic analysis. Dev. Biol. 134, 189–200. Abstract Article
Skehel, P.A., Fabian-Fine, R., and Kandel, E.R. (2000). Mouse VAP33 is associated with the endoplasmic reticulum and microtubules. Proc. Natl. Acad. Sci. USA 97, 1101–1106. Abstract Article
Skehel, P.A., Martin, K.C., Kandel, E.R., and Bartsch, D. (1995). A VAMP-binding protein from Aplysia required for neurotransmitter release. Science 269, 1580–1583. Abstract
Smith, H.E., and Ward, S. (1998). Identification of protein-protein interactions of the major sperm protein (MSP) of Caenorhabditis elegans. J. Mol. Biol. 279, 605–619. Abstract Article
Ward, S. (1987). Expression of sperm-specific genes during nematode spermatogenesis. Ann. N.Y. Acad. Sci. 513, 128–133. Abstract
Ward, S., Argon, Y., and Nelson, G.A. (1981). Sperm morphogenesis in wild-type and fertilization-defective mutants of Caenorhabditis elegans. J. Cell Biol. 91, 26–44. Abstract Article
Ward, S., Burke, D.J., Sulston, J.E., Coulson, A.R., Albertson, D.G., Ammons, D., Klass, M., and Hogan, E. (1988). Genomic organization of major sperm protein genes and pseudogenes in the nematode Caenorhabditis elegans. J. Mol. Biol. 199, 1–13. Abstract Article
Ward, S., and Carrel, J.S. (1979). Fertilization and sperm competition in the nematode Caenorhabditis elegans. Dev. Biol. 73, 304–321. Abstract
*Edited by Donald G. Moerman and James M. Kramer. Last revised April 21, 2005. Published February 1, 2006. This chapter should be cited as: Smith, H. Sperm motility and MSP (February 1, 2006), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.68.1, http://www.wormbook.org.
Copyright: © 2006 Harold Smith. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
§To whom correspondence should be addressed. E-mail: smithh@umbi.umd.edu
 All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.
All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.