Embryo series courtesy of Einhard Schierenberg
Embryo series courtesy of Einhard SchierenbergTable of Contents
Abstract
TGF-β superfamily ligands play fundamental roles in the development and physiology of diverse animal species. Genetic and genomic analyses in the model organism Caenorhabditis elegans have contributed to the understanding of TGF-β-related signal transduction mechanisms. In this chapter, I describe the currently characterized TGF-β-related signals and signal transduction cassettes in C. elegans. Homology searches of the genome identify five TGF-β-related genes, for which functions have been identified for three. Two of the TGF-β-related genes, daf-7 and dbl-1, function through conventional signaling pathways. These signaling pathways are comprised of ser/thr kinase receptors, Smads, and transcription co-factors. A third TGF-β-related gene, unc-129, functions in axonal guidance using novel signaling mechanisms. Thus, TGF-β-related signaling in C. elegans proceeds via both conserved and novel paradigms that can inform studies in other animal systems.
TGF-β is the prototype of a large family of secreted peptide growth factors in metazoans that includes activin and the Bone Morphogenetic Proteins (BMPs) in vertebrates. The canonical TGF-β signal transduction pathway (ten Dijke and Hill, 2004) is comprised of two transmembrane ser/thr kinase receptors (type I and type II) and two or three intracellular Smad signal transducers. Upon ligand binding, the type I and type II receptors are recruited into a heteromeric complex and the type II receptor phosphorylates the type I receptor on the conserved GS domain. The type I receptor kinase is thereby activated, and phosphorylates Smads of the R-Smad (receptor-regulated) subfamily. R-Smads form heteromeric complexes with Co-Smads and accumulate in the nucleus to regulate gene transcription. A third group of Smads, I-Smads, act as inhibitors of pathway signaling by interfering with productive molecular interactions. Smads contain two regions of homology, the N-terminal MH1 domain that binds DNA and the C-terminal MH2 domain that participates in intermolecular interactions and activates transcription.
In C. elegans, five TGF-β-related genes can be identified by sequence homology: daf-7, dbl-1, unc-129, tig-2, and Y46E12BL.1 (Table 1). No biological roles have yet been described for tig-2 or Y46E12BL.1. The unc-129 signaling pathway is still being elucidated, but it does not seem to depend on known TGF-β-related receptors or Smads. Conventional signaling pathways have been identified that act downstream of daf-7 and dbl-1, and are referred to as the Dauer pathway (Figure 1) and the Sma/Mab pathway (Figure 2), respectively. Most of the components of these two pathways were identified by forward genetic approaches, and often the signal transduction components were identified before the relevant ligands. More recent work using reverse genetics, yeast two-hybrid, and DNA microarray analysis continues to identify components, modifiers, and targets of these pathways and to uncover new biological roles for existing pathways and components. A summary of the TGF-β signaling components in C. elegans and their likely Drosophila and mammalian orthologs is given in Table 1. In addition to the genes described below, there is one potential I-Smad gene in the genome, tag-68 (Newfeld et al., 1999), whose function has not yet been determined.
Table 1. TGF-β signaling components in C. elegans
| Component | Gene name | Drosophila ortholog | Mammalian ortholog |
|---|---|---|---|
| Ligand | daf-7 | Alp23B | GDF11 |
| dbl-1 | dpp | BMP5 | |
| unc-129 | – | – | |
| tig-2 | gbb | BMP8 | |
| Y46E12BL.1 | – | – | |
| Type I Receptor | daf-1 | babo | TGF-βRI |
| sma-6 | tkv | BMPRIB | |
| Type II Receptor | daf-4 | put | ACTRIIB |
| R-Smad | daf-8 | Smox | – |
| daf-14 | Similar to Smox | Smad2 | |
| sma-2 | Mad | Smad1 | |
| sma-3 | Similar to Mad | Smad5 | |
| Co-Smad | daf-3 | – | – |
| sma-4 | Medea | Smad4 | |
| I-Smad | tag-68 | Similar to Dad | Similar to Smad6 |
| Transcription factors | daf-5 | – | Similar to Sno/Ski |
| daf-12 | Hr96 (nuclear hormone receptor) | – | |
| sma-9 | shn (Zn finger protein) | Similar to HIVEP1 | |
| lin-31 | fd95Cb | FKH-5 |
The dauer larva is an alternative third larval stage specialized for survival under harsh environmental conditions (high population density, low food availability, high temperature; Cassada and Russell, 1975; Golden and Riddle, 1984a; Golden and Riddle, 1984b). The decision to enter the dauer stage is made in the L1 and L2 stages. When dauer larvae encounter more favorable environments, they exit dauer and resume development, entering the L4 stage. Large-scale genetic screens were conducted for mutations that affect the decision to enter dauer (Riddle et al., 1981; Golden and Riddle 1984a; Malone and Thomas, 1994). The mutants isolated are either dauer-constitutive (Daf-c: entering dauer inappropriately under favorable environmental conditions) or dauer-defective (Daf-d: failing to enter dauer under adverse conditions). Genetic epistasis and detailed phenotypic analysis allowed these genes to be grouped into three classes (Thomas et al., 1993). Subsequent molecular analysis established that the genes in each class encode components of three different signaling pathways: a TGF-β-related pathway, an insulin-related pathway, and a cyclic nucleotide pathway. In this section, I will refer to the TGF-β-related pathway that functions in dauer development as the Dauer TGF-β pathway. See appropriate chapters for more information on dauer development and the insulin and cyclic nucleotide signaling pathways.
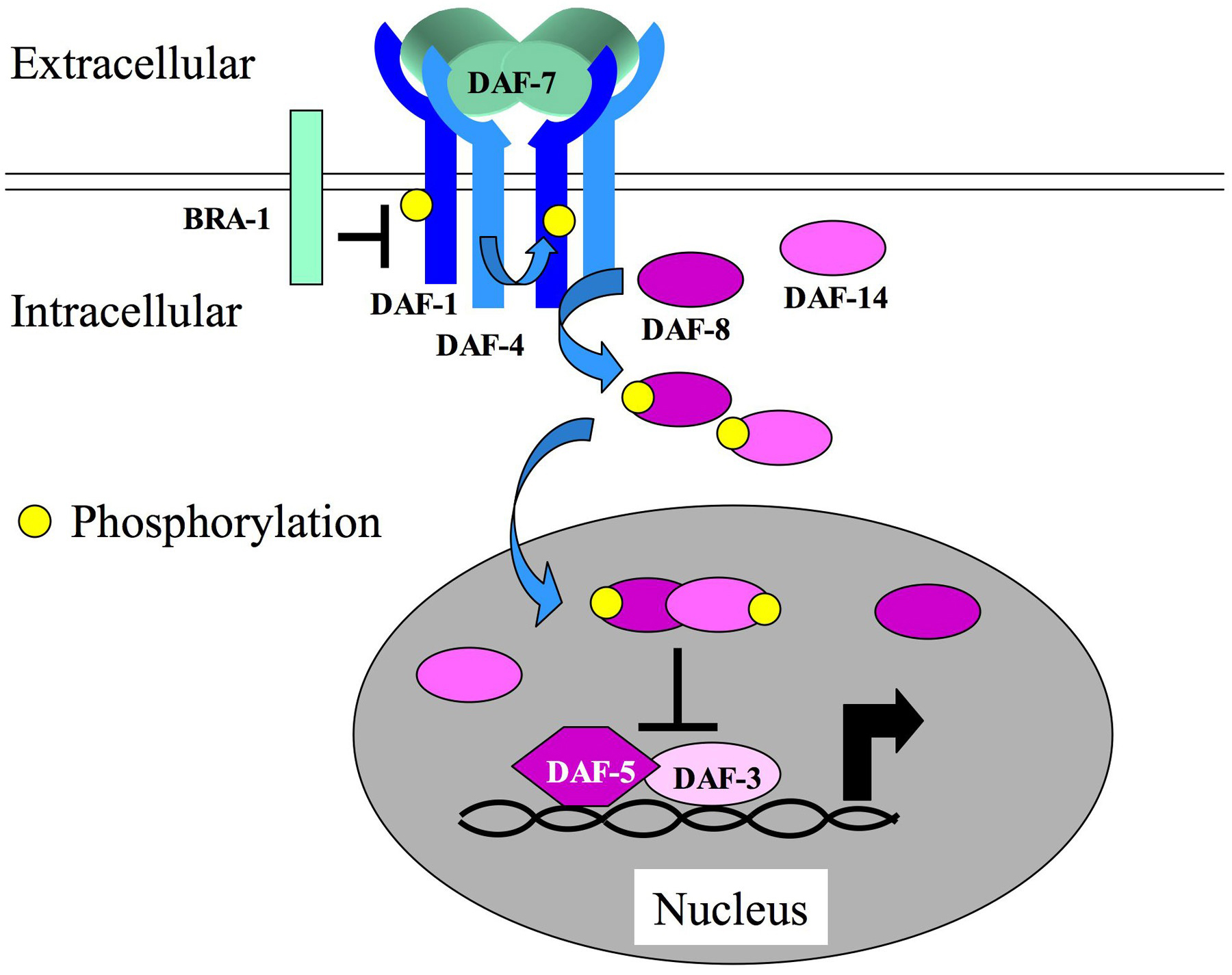
DAF-7 is the TGF-β-related ligand for the Dauer pathway (Figure 1; Ren et al., 1996). daf-7 mutants are Daf-c, indicating that the gene has a role in promoting continuous, non-Dauer development. DAF-7 is expressed in the pair of ASI sensory neurons under favorable environmental conditions (Ren et al., 1996; Schackwitz et al., 1996). Its expression is repressed under dauer-inducing conditions, leading to a loss of Dauer pathway signaling and entry into the dauer stage. Four other Daf-c genes are characterized by alleles that confer phenotypes, and have genetic interactions similar to those of mutations in daf-7, suggesting that they function with daf-7. These are daf-1, daf-4, daf-8, and daf-14. Genetic epistasis analysis has predicted that three Daf-d genes act downstream of this pathway: daf-3, daf-5, and daf-12 (Thomas et al., 1993). Mutations in any of these three Daf-d genes suppress the effects of mutations in daf-7, daf-1, daf-4, daf-8 or daf-14. daf-3 and daf-5 seem to function specifically in this pathway since they do not strongly suppress mutations in other Daf-c genes. In contrast, daf-12 can suppress mutations both in daf-7 class genes and in other Daf-c genes, suggesting that it integrates signals from multiple pathways. In addition to the Daf-c phenotype, DAF-7 pathway genes also have a less well-characterized role in egg laying. The egg-laying defect is suppressed by mutations in daf-3 and daf-5 (Trent et al., 1983). Mutations in two other genes required for egg laying, egl-4 and egl-32, are also suppressed by daf-3 and daf-5 mutations, indicating a possible interaction with this pathway. egl-4 has additional phenotypes, suggesting roles in body size regulation (see below) and chemosensation, and encodes multiple cGMP-dependent protein kinase isoforms (Fujiwara et al., 2002; L′Etoile et al., 2002; Hirose et al., 2003).
Molecular cloning of the daf-7 class genes has revealed that they comprise a TGF-β signaling pathway. daf-1 and daf-4 encode type I and type II TGF-β receptors, respectively (Georgi et al., 1990; Estevez et al., 1993). The molecular cloning of daf-4 was pivotal in the recognition of this pathway as a TGF-β signaling pathway, since DAF-4 showed homology to the newly identified TGF-β type II receptors. (The cloning of daf-1 preceded this time, so its homology was not recognized until later). Importantly, DAF-4 was shown to bind human BMP ligands in cell culture, illustrating the conservation of these pathway components from nematodes to humans (Estevez et al., 1993). daf-4 is the sole type II receptor gene in C. elegans; it functions both in the Dauer pathway and in the Sma/Mab pathway by interacting with different pathway-specific components. daf-4 is expressed widely during all stages of C. elegans development (Patterson et al., 1997; Gunther et al., 2000). Alternative polyadenylation of the daf-4 locus produces a truncated protein containing only the extracellular domain that acts as a negative regulator of DAF-7 function (Gunther and Riddle, 2004). daf-1, the type I receptor, is also widely expressed, primarily in the nervous system (Gunther et al., 2000). Genetic evidence suggests that DAF-1 is capable of signaling, albeit weakly, in the absence of phosphorylation by DAF-4. Thus, in a daf-4 mutant background, depletion of daf-1 causes a more severe mutant phenotype and overexpression of daf-1 partially rescues the mutant phenotype. The ability of DAF-1 to be constitutively active is supported by the phylogeny of TGF-β receptors that indicates that DAF-1 is somewhat related to both type I and type II receptors (Newfeld et al., 1999). BRA-1 is a DAF-1 binding protein identified by homology to human BMP receptor-associated molecule BRAM1, which negatively regulates DAF-7 signaling (Morita et al., 2001).
|
Figure 1. The Dauer TGF-β-related pathway . DAF-7 promotes continuous, nonDauer development. The DAF-7 signal is transduced by DAF-1 type I receptor, DAF-4 type II receptor, and DAF-8 and DAF-14 Smads. These components, when activated, inhibit the functions of DAF-3 Smad and DAF-5 Sno/Ski, which promote Dauer development. BRA-1 is a negative regulator of DAF-1.
The Smads participating in the Dauer pathway are encoded by daf-8, daf-14, and daf-3. DAF-8 and DAF-14 are related to R-Smads, except that DAF-14 lacks strong homology to the MH1 domain (Estevez 1997; Inoue and Thomas, 2000). These R-Smads have been shown to have partially redundant functions (Inoue and Thomas, 2000). daf-3 encodes a Smad distantly related to Co-Smads (Patterson et al., 1997). All of these Smads are expressed broadly. While DAF-8 and DAF-14 function to transduce the daf-7 signal, DAF-3 activity is negatively regulated by the pathway. In the absence of DAF-3, such as in a daf-3 mutant, animals enter the non-Dauer L3 stage regardless of the presence or absence of DAF-7 signaling (Patterson et al.1997). Thus, the output of Dauer TGF-β pathway signaling culminates in the inhibition of daf-3 activity and is dispensable in the absence of daf-3. This type of antagonistic interaction differs from the mechanism of inhibition by I-Smads in other systems. daf-5 has the same type of genetic interactions with the Dauer TGFβ pathway as daf-3. Cloning of daf-5 revealed homology to the Sno/Ski family of oncogenes that negatively regulate TGF-β signaling in vertebrates (da Graca et al., 2004; Tewari et al., 2004). DAF-5 physically interacts with DAF-3 to promote dauer development. Finally, like daf-3 and daf-5 mutations, daf-12 mutations suppress the Daf-c phenotype of Dauer pathway mutants. Unlike daf-3 and daf-5, however, daf-12 also shows strong suppression of mutations in the insulin-related and cyclic nucleotide pathways. daf-12 encodes a nuclear hormone receptor that is required for regulating multiple stage-specific aspects of nematode development (Antebi et al., 2000; Snow and Larsen, 2000). The daf-12 locus is complex, encoding multiple alternatively spliced isoforms, and is capable of mutating to Daf-d, Daf-c, and heterochronic phenotypes independently.
In large-scale interactome mapping analysis of this pathway using the yeast two-hybrid system, physical interactions among 59 proteins, including DAF-1 type I receptor, DAF-4 type II receptor, DAF-3 Smad, DAF-5 Sno/Ski, and several novel interactors were identified (Tewari et al., 2004). In these screens, a physical interaction between DAF-3 and DAF-5 was shown. Among the novel interactors was DAF-21/HSP90, which was previously shown to function in the cyclic nucleotide pathway and now found to interact physically with DAF-1 and DAF-4. TTX-1, a homeodomain protein required for thermosensation, is linked with both DAF-3 Smad and DAF-4 type II receptor indirectly via secondary interactions. DAF-3 Smad also shows direct or indirect associations with SWI3 and SNF5 homologs (Y11G7B.23 and R07E5.3, respectively).
The Sma/Mab pathway regulates body size and the development of male-specific sensory rays and copulatory spicules (Savage et al., 1996). Thus, mutants of the pathway are small in size (Small) and mutant males have patterning defects in the male tail (Male abnormal). The normal male tail contains nine bilaterally symmetrical pairs of sensory organs, known as rays, embedded in a cuticular fan (Sulston et al., 1980). In Sma/Mab pathway mutants, defects in the specification of ray identities lead to the fusion of pairs of rays (4-5, 6-7, and 8-9) with varying penetrance. The defects in ray identity specification are accompanied by changes in neurotransmitter expression by ray neurons (Lints and Emmons, 1999). Furthermore, in mutant males, the copulatory spicules are crumpled. The Mab defects prevent homozygous mutant males from mating successfully. For more information on male tail development and function, see chapters on Male development and Male mating behavior.
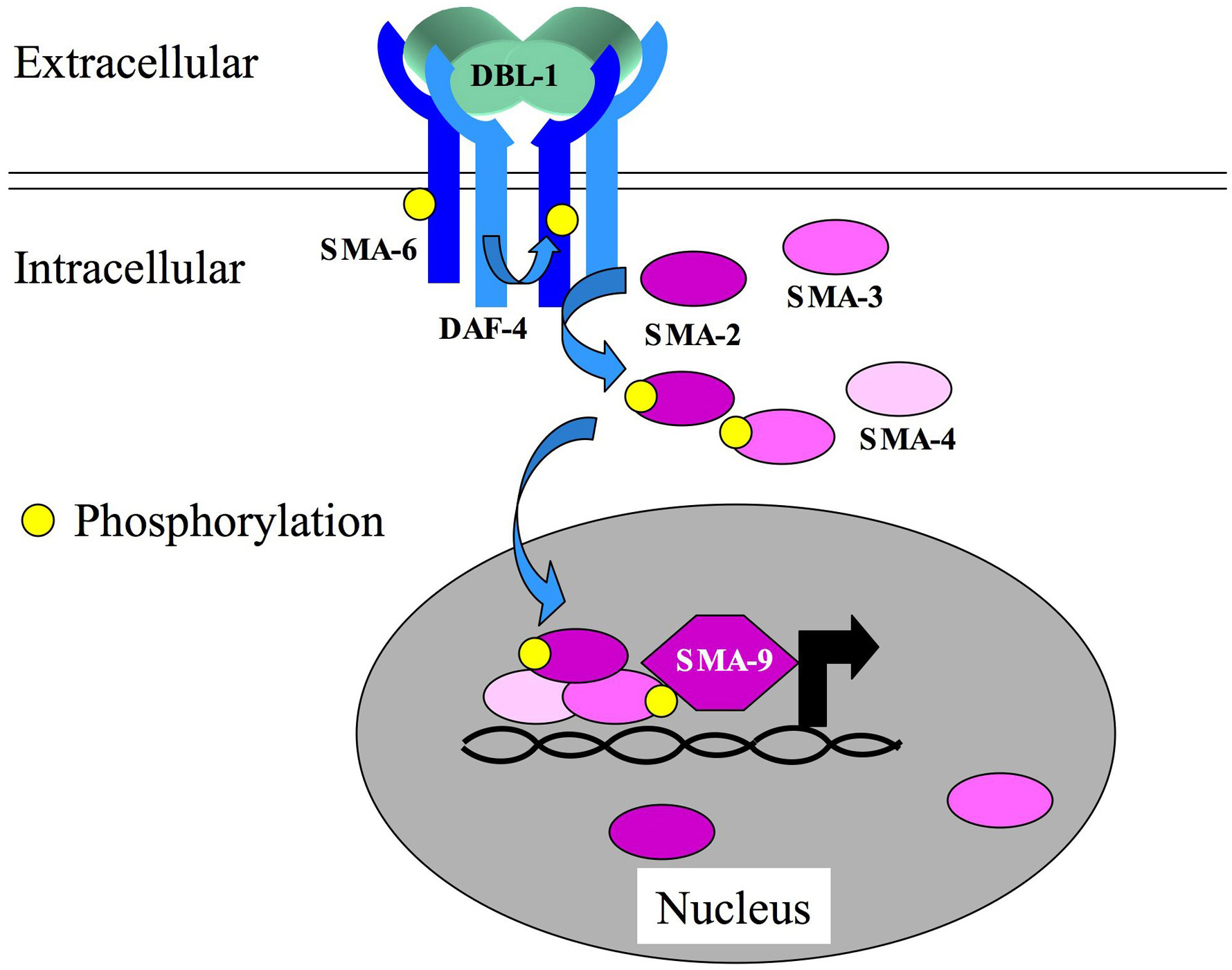
DBL-1 is the TGF-β-related ligand for the Sma/Mab pathway (Figure 2; Morita et al., 1999; Suzuki et al., 1999). dbl-1 mutations were generated using two strategies: (1) reverse genetics, by deletion of dbl-1 (DPP/BMP-like) sequences (Morita et al., 1999; Suzuki et al., 1999); and (2) forward genetics, by isolation of Sma mutants and subsequent mapping and complementation tests (Suzuki et al., 1999); Savage-Dunn et al., 2003). DBL-1 acts as a dose-dependent regulator of body size (Morita et al., 1999; Suzuki et al., 1999). Loss of dbl-1 activity results in smaller animals, while dbl-1 overexpression results in longer animals. In dbl-1 mutant males, the spicules are crumpled and there are defects in the specification of rays 5, 7, and 9. Overexpression of dbl-1 in males leads to fusions of rays 3 and 4 (Suzuki et al., 1999). dbl-1 is expressed primarily in neurons in both hermaphrodites and males (Morita et al., 1999; Suzuki et al., 1999). Although overexpression of dbl-1 leads to gain-of-function phenotypes, overexpression of the downstream signaling components does not, suggesting that the concentration of ligand is limiting.
Candidate signaling components of the Sma/Mab pathway were identified by the study of mutants with similar Sma and Mab phenotypes. Some of these mutants had existed for many years in strain collections (sma-2, sma-3, sma-4, sma-6), while others were identified by reverse genetics and additional genetic screens (as with dbl-1 itself and sma-9). The receptors for DBL-1 are encoded by sma-6 (type I), which is pathway-specific (Krishna et al., 1999), and daf-4 (type II), which also functions in the Dauer pathway (Estevez et al., 1993). The signal transducers for this pathway are SMA-2, SMA-3, and SMA-4 (Savage et al., 1996), which, together with Drosophila Mad, were the founding members of the Smad family. SMA-2 and SMA-3 are R-Smad homologs, and SMA-4 is a Co-Smad homolog. Unlike the partial redundancy between DAF-8 and DAF-14, R-Smads, no function of either SMA-2 or SMA-3 is detectable in the absence of the other partner, or of SMA-4 Co-Smad (Savage-Dunn et al., 2000). This observation has led to the suggestion that the functional Smad complex for this pathway is a heterotrimer (SMA-2/SMA-3/SMA-4). While early models of TGF-β signaling suggested that R-Smad phosphorylation and heteromeric complex formation were necessary for nuclear translocation, it has more recently become evident that Smads engage in dynamic nucleocytoplasmic shuttling (Xiao et al., 2001; Inman et al., 2002; Xu et al., 2002; Nicolas et al., 2004). Similarly, a rescuing SMA-3::GFP fusion protein is found in both the cytoplasm and the nucleus (Wang et al., 2002). In wild-type animals, the protein accumulates to high levels in the nucleus, while in sma-6 type I receptor mutants, the protein is more evenly partitioned. Nuclear accumulation is not influenced by R-Smad (sma-2) or Co-Smad (sma-4) depletion.
Expression patterns reported for the receptors and Smads are similar and include expression in the pharynx, intestine, and hypodermis. Genetic mosaic analyses and directed expression studies of daf-4, sma-6, and sma-3 have indicated that the expression of signaling components in the hypodermis is necessary and sufficient for body size regulation (Inoue and Thomas, 2000; Yoshida et al., 2001; Wang et al., 2002). The expression in the pharynx and intestine may reflect a more recently uncovered role for the Sma/Mab pathway in innate immunity (Mallo et al., 2002; see Signaling in the immune response). Some of the transcriptional target genes of this pathway were identified by differential hybridization of a cDNA array (Mochii et al., 1999). One of these target genes is sma-6, indicating the existence of a positive feedback loop. Some of the other target genes were expressed in the intestine and had functional domains, such as C-type lectin domains, suggestive of a role in immune recognition of pathogens. In addition, the Hox gene ceh-13, a labial homolog, has been identified as a direct or indirect target of Sma/Mab signaling in the male tail (Stoyanov et al., 2003). ceh-13 expression in the B cell lineage and in rays 5, 7, and 9 is dependent on the function of the Sma/Mab pathway.
SMA-9 is a transcriptional co-factor for the Sma/Mab pathway (Liang et al., 2003). Identified in a genetic screen for small mutants (Savage-Dunn et al., 2003), sma-9 encodes a large zinc finger transcription factor related to Drosophila Schnurri (Liang et al., 2003). Phenotypic analysis of sma-9 indicates that it functions in a subset of Sma/Mab signaling outcomes: early but not late larval growth; ray 9 but not ray 5 and 7 morphogenesis; regulation of neurotransmitter expression in rays 5, 7, and 9; and spicule morphogenesis with reduced penetrance. This observation suggests that SMA-9 may be required for transcriptional regulation of a subset of Sma/Mab pathway target genes. SMA-9 is nuclear localized and detected in most, if not all, cells by immunocytochemistry. The sma-9 locus shows extensive alternative splicing that could give rise to isoforms with different numbers of zinc finger motifs and trans splicing that could produce shorter N-terminal and C-terminal isoforms. Another transcriptional co-factor that may be involved in a tissue-specific output of this pathway is the forkhead transcription factor LIN-31. lin-31 was originally identified by its role in vulval development (Miller et al., 1993), but lin-31 mutant males also have a crumpled spicule defect like that of Sma/Mab pathway mutants (Baird and Ellazar, 1999). Since forkhead transcription factors in other systems function as Smad co-factors (Chen et al., 1996), LIN-31 may similarly interact with SMA-2/SMA-3/SMA-4 specifically to regulate spicule morphogenesis.
rnt-1 encodes a runt domain transcription factor that may function in parallel with the Sma/Mab pathway in regulation of body size and male tail morphogenesis (Ji et al., 2004). rnt-1 was identified by its homology to mammalian RUNX transcription factors (Bae and Lee, 2000), which interact with Smads (Hanai et al., 1999; Zhang et al., 2000; Alliston et al., 2001). A rnt-1 deletion mutant is small and double mutants with rnt-1 and sma-2, sma-3, sma-4, or sma-6 are smaller than any of the single mutants (Ji et al., 2004). rnt-1 is expressed in the hypodermal seam cells and the intestine (Nam et al., 2002). rnt-1 mutants also have defects in the male tail, but these are more severe and less specific than the Sma/Mab pathway defects, indicating that rnt-1 has additional functions outside of this pathway (Ji et al., 2004). RNT-1 physically interacts with SMA-4 Smad, providing a possible mechanism for the synergistic interactions suggested by the mutant phenotypes.
Body size and morphology are regulated by several different mechanisms in C. elegans. Dumpy (Dpy) mutants are often defective in cuticle collagen genes (Johnstone, 2000) or dosage compensation genes (Plenefisch et al., 1989). A smaller number of genes can mutate to result in long (Lon) animals. Sma mutants were originally identified by Sydney Brenner by their relatively proportional reduction in size (Brenner, 1974). Most of these Sma mutants are defective in components of the TGF-β Sma/Mab pathway, indicating that this pathway is a major regulator of body size in C. elegans. In other animals, components of insulin signaling pathways and TOR kinase signaling regulate cell size (Schmelzle and Hall, 2000; Oldham and Hafen, 2003). In C. elegans, however, mutations in these components have not been shown to affect body size but rather play a role in dauer development (Kimura et al., 1997; Jia et al., 2004).
Mutants in the Sma/Mab pathway hatch as L1 larvae indistinguishable in size from wild type (Suzuki et al., 1999; Savage-Dunn et al., 2000). Postembryonic growth is then reduced, resulting in a reduction in both body length and volume (Savage-Dunn et al., 2000; Hirose et al., 2003). Sma/Mab mutant adults contain the same number of nuclei as wild-type adults, indicating that cell size must be smaller (Nagamatsu and Ohshima, 2004). Despite the relatively proportional appearance of Sma/Mab mutants, organ size measurements have shown that different organs are reduced in size to different degrees. The degree of reduction in the size of hypodermal seam cells and hyp7 is most proportional to the degree of reduction in body size (Wang et al., 2002; Nagamatsu and Ohshima, 2004), consistent with the defined focus of action of Sma/Mab signaling components in the hypodermis. In contrast to these results, sma-1 βH-spectrin mutants have a defect in embryonic elongation, but not postembryonic growth, and organ size measurements reveal a more severe reduction in pharynx length than in seam cell length (Wang et al., 2002). The molecular mechanisms of cell size control by these genes remain elusive. A correlation between body size and hypodermal ploidy has been described for some mutants (Flemming et al., 2000; Morita et al., 2002; Nystrom et al., 2002), but may not be sufficient to account for all of the differences in size seen in mutant animals (Nagamatsu and Ohshima, 2004; Nystrom et al., 2002).
Several genes can mutate to a Lon phenotype similar to dbl-1 overexpression, resulting in an increase in body length but not in volume. lon-1 is a putative target of the Sma/Mab pathway and encodes a member of the CRISP (cysteine-rich secretory protein) family (Maduzia et al., 2002; Morita et al., 2002). Mutations in lon-1 suppress the small body size phenotype of Sma/Mab pathway mutants, suggesting that lon-1 acts downstream. lon-3 is a cuticle collagen gene for which loss-of-function mutants are long but overexpressing animals are small (Nystrom et al., 2002; Suzuki et al., 2002). The Lon phenotype of lon-3, together with the Dpy phenotypes of other cuticle collagen genes, indicates a role for cuticle collagens in constraining the body size and shape of C. elegans. In addition to these long mutants, large body size mutants with increased body volume were sought in a genetic screen (Hirose et al., 2003). In this screen, multiple alleles of the gene egl-4 were isolated. egl-4, which encodes cGMP-dependent protein kinases, has additional phenotypes indicating roles in egg laying (see above) and sensory processing (Daniels et al., 2000; Fujiwara et al., 2002; L′Etoile et al., 2002). In egl-4 mutants, most of the major organs except the gonad are increased in volume without an increase in the number of nuclei (Hirose et al., 2003) and there is evidence of an increase in fluid retention (Nagamatsu and Ohshima, 2004). The gene is expressed in neurons, hypodermis, intestine, and body wall muscles (Fujiwara et al., 2002; L′Etoile et al., 2002; Hirose et al., 2003). Expression in neurons or in hypodermis can rescue the organ size defects nonautonomously (Nakano et al., 2004). Mutations in Sma/Mab pathway components can completely suppress the large body size phenotype of egl-4, suggesting that egl-4 acts upstream of the TGF-β pathway (Hirose et al., 2003).
unc-129, a TGF-β-related gene (Colavita et al., 1998), was isolated in a genetic screen (Colavita and Culotti, 1998) designed to identify genes that interact with unc-5, which encodes a transmembrane receptor in the UNC-6/netrin pathway necessary for axon guidance along the dorsoventral axis. unc-129 mutants, like unc-5, unc-6, and unc-40 mutants, have defects in the trajectories of some dorsally migrating motoraxons (Colavita and Culotti, 1998). In addition, both unc-129 and dbl-1 mutations can enhance a defect in the migration of the gonadal distal tip cells in a weak unc-5 mutant background (Merz et al., 2003). UNC-129 is expressed in several tissues including dorsal body wall muscle, in which its expression is sufficient to provide normal axon guidance cues (Colavita et al., 1998). UNC-129 is unlikely to interact with the identified TGF-β superfamily receptors DAF-4, DAF-1, or SMA-6, since mutations in the receptor genes do not cause defects in axon guidance. In addition to its axon guidance role, a redundant role for unc-129 in male tail sensory ray development has been identified (Ikegami et al., 2004). In this pathway, unc-129 interacts with the semaphorin gene, mab-20 and the plexin gene, plx-2, and acts in parallel with semaphorin/mab-20 and ephrin/efn-4, to prevent sensory ray fusions between adjacent rays. Interestingly, unc-129 is expressed in alternating rays (1, 3, 5, and 7), possibly to keep their identities distinct from those of their neighbors.
Work in the author's laboratory is supported in part by Research Scholar Grant 98-230-04-DDC from the American Cancer Society. I thank Jun Liang and Marie McGovern forcritical reading of the manuscript.
Alliston, T., Choy, L., Ducy, P., Karsenty, G., and Derynck, R. (2001). TGF-β induced repression of CBFA1 by Smad3 decreases cbfa1 and osteocalcin expression and inhibits osteoblast differentiation. EMBO J. 20, 2254–2272. Abstract Article
Antebi, A., Yeh, W.H., Tait, D., Hedgecock, E.M., and Riddle, D.L. (2000). daf-12 encodes a nuclear receptor that regulates the dauer diapause and developmental age in C. elegans. Genes Dev. 14, 1512–1527. Abstract
Bae, S.C., and Lee, J. (2000). cDNA cloning of run, a Caenorhabditis elegans Runt domain encoding gene. Gene 241, 255–258. Abstract Article
Baird, S.E., and Ellazar, S.A. (1999). TGFb-like signaling and spicule development in Caenorhabditis elegans. Dev. Biol. 212, 93–100. Abstract Article
Brenner, S. (1974). The genetics of Caenorhabditis elegans. Genetics 77, 71–94. Abstract
Cassada, R.C., and Russell, R.L. (1975). The dauer larva, a post-embryonic developmental variant of the nematode Caenorhabditis elegans. Dev. Biol. 46, 326–342. Abstract Article
Chen, X., Rubock, M.J., and Whitman, M. (1996). A transcriptional partner for MAD proteins in TGF-β signalling. Nature 383, 691–696. Abstract Article
Colavita, A., and Culotti, J.G. (1998). Suppressors of ectopic UNC-5 growth cone steering identify eight genes involved in axon guidance in Caenorhabditis elegans. Dev. Biol. 194, 72–85. Abstract Article
Colavita, A., Krishna, S., Zheng, H., Padgett, R.W., and Culotti, J.G. (1998). Pioneer axon guidance by UNC-129, a C. elegans TGF-β. Science 281, 706–709. Abstract Article
da Graca, L.S., Zimmerman, K.K., Mitchell, M.C., Kozhan-Gorodetska, M., Sekiewica, K., Morales, Y., and Patterson, G.I. (2004). DAF-5 is a Ski oncoprotein homolog that functions in a neuronal TGFb pathway to regulate C. elegans dauer development. Development 131, 435–446. Abstract Article
Daniels, S.A., Ailion, M., Thomas, J.H., and Sengupta, P. (2000). egl-4 acts through a transforming growth factor-b/SMAD pathway in Caenorhabditis elegans to regulate multiple neuronal circuits in response to sensory cues. Genetics 156, 123–141. Abstract
Estevez, A.O.Z. (1997). The role of the daf-8 gene in Caenorhabditis elegans dauer larva development. PhD thesis, University of Missouri, Columbia, MO.
Estevez, M., Attisano, L., Wrana, J.L., Albert, P.S., Massague, J., and Riddle, D.L. (1993). The daf-4 gene encodes a bone morphogenetic protein receptor controlling C. elegans dauer larva development. Nature 365, 644–649. Abstract Article
Flemming, A.J., Shen, Z.Z., Cunha, A., Emmons, S.W., and Leroi, A.M. (2000). Somatic polyploidization and cellular proliferation drive body size evolution in nematodes. Proc. Natl. Acad. Sci. USA 97, 5285–5290. Abstract Article
Fujiwara, M., Sengupta, P., and McIntire, S.L. (2002). Regulation of body size and behavioral state of C. elegans by sensory perception and the EGL-4 cGMP-dependent protein kinase. Neuron 36, 1091–1102. Abstract Article
Georgi, L.L., Albert, P.S., and Riddle, D.L. (1990). daf-1, a C. elegans gene controlling dauer larva development, encodes a novel receptor protein kinase. Cell 61, 635–645. Abstract Article
Golden, J.W., and Riddle, D.L. (1984a). A pheromone-induced developmental switch in Caenorhabditis elegans : temperature-sensitive mutants reveal a wild-type temperature-dependent process. Proc. Natl. Acad. Sci. USA 81, 819–823. Abstract
Golden, J.W., and Riddle, D.L. (1984b). The Caenorhabditis elegans dauer larva: developmental effects of pheromone, food, and temperature. Dev. Biol. 102, 368–378. Abstract Article
Gunther, C.V., Georgi, L.L., and Riddle, D.L. (2000). A Caenorhabditis elegans type I TGFb receptor can function in the absence of type II kinase to promoter larval development. Development 127, 3337–3347 Abstract
Gunther, C.V., and Riddle, D.L. (2004). Alternative polyadenylation results in a truncated daf-4 BMP receptor that antagonizes DAF-7-mediated development in C. elegans. J. Biol. Chem. 279, 39555–39565. Abstract Article
Hanai, J., Chen, L.F., Kanno, T., Ohtani-Fujita, N., Kim, W.Y., Guo, W.H., Imamura, T., Ishidou, Y., Fukuchi, M., Shi, M.J., et al. (1999). Interaction and functional cooperation of PEBP2/CBF with Smads. Synergistic induction of the immunoglobulin germline Ca promoter. J. Biol. Chem. 274, 31577–31582. Abstract Article
Hirose, T., Nakano, Y., Nagamatsu, Y., Misumi, T., Ohta, H., and Ohshima, Y. (2003). Cyclic GMP-dependent protein kinase EGL-4 controls body size and lifespan in C. elegans. Development 130, 1089–1099. Abstract Article
Ikegami, R., Zheng, H., Ong, S.H., and Culotti, J. (2004). Integration of semaphorin-2A/MAB-20, ephrin-4, and UNC-129 TGF-β signaling pathways regulates sorting of distinct sensory rays in C. elegans. Dev. Cell 6, 383–395. Abstract Article
Inman, G.J., Nicolas, F.J., and Hill, C.S. (2002). Nucleocytoplasmic shuttling of Smads 2, 3, and 4 permits sensing of TGF-β receptor activity. Mol. Cell 10, 283–294. Abstract Article
Inoue, T., and Thomas, J.H. (2000). Targets of TGF-β signaling in Caenorhabditis elegans dauer formation. Dev. Biol. 217, 192–204. Abstract Article
Ji, Y.J., Nam, S., Jin, Y.H., Cha, E.J., Lee, K.S., Choi, K.Y., Song, H.O., Lee, J., Bae, S.C., and Ahnn, J. (2004). RNT-1, the C. elegans homologue of mammalian RUNX transcription factors, regulates body size and male tail development. Dev. Biol. 274, 402–412. Abstract Article
Jia, K., Chen, D., and Riddle, D.L. (2004). The TOR pathway interacts with the insulin signaling pathway to regulate C. elegans larval development, metabolism and life span. Development 131, 3897–3906. Abstract Article
Johnstone, I.L. (2000). Cuticle collagen genes. Expression in Caenorhabditis elegans. Trends Genet. 16, 21–27. Abstract Article
Kimura, K.D., Tissenbaum, H.A., Liu, Y., and Ruvkun, G. (1997). daf-2, an insulin receptor-like gene that regulates longevity and diapause in Caenorhabditis elegans. Science 277, 942–946. Abstract Article
Krishna, S., Maduzia, L.L., and Padgett, R.W. (1999). Specificity of TGFb signaling is conferred by distinct type I receptors and their associated SMAD proteins in Caenorhabditis elegans. Development 126, 251–260. Abstract
L'Etoile, N.D., Coburn, C.M., Eastham, J., Kistler, A., Gallegos, G., and Bargmann, C.I. (2002). The cyclic GMP-dependent protein kinase EGL-4 lates olfactory adaptation in C. elegans. Neuron 36, 1079–1089. Abstract Article
Liang, J., Lints, R., Foehr, M.L., Tokarz, R., Yu, L., Emmons, S.W., Liu, J., and Savage-Dunn, C. (2003). The Caenorhabditis elegans schnurri homolog sma-9 mediates stage- and cell type-specific responses to DBL-1 BMP-related signaling. Development 130, 6453–6464. Abstract Article
Lints, R., and Emmons, S.W. (1999). Patterning of dopaminergic neurotransmitter identity among Caenorhabditis elegans ray sensory neurons by a family signaling pathway and a Hox gene. Development 126, 5819–5831. Abstract
Maduzia, L.L, Gumienny, T.L., Zimmerman, C.M., Wang, H., Shetgiri, P., Krishna, S., Roberts, A.F., and Padgett, R.W. (2002). lon-1 regulates Caenorhabditis elegans body size downstream of the dbl-1 TGFb signaling pathway. Dev. Biol. 246, 418–428. Abstract Article
Mallo, G.V., Kurz, C.L., Couillault, C., Pujol, N., Granjeaud, S., Kohara, Y., and Ewbank, J.J. (2002). Inducible antibacterial defense system in C.elegans. Curr. Biol. 12, 1209–1214. Abstract Article
Malone, E.A., and Thomas, J.H. (1994). A screen for nonconditional dauer-constitutive mutations in Caenorhabditis elegans. Genetics 136, 879–886. Abstract
Merz, D.C., Alves, G., Kawano, T., Zheng, H., and Culotti, J.G. (2003). UNC-52/perlecan affects gonadal leader cell migrations in C. elegans hermaphrodites through alterations in growth factor signaling. Dev. Biol. 256, 173–186. Abstract Article
Miller, L.M., Gallegos, M.E., Morisseau, B.A., and Kim, S.K. (1993). lin-31, a Caenorhabditis elegans HNF-3/fork head transcription factor homolog, specifiies three alternative cell fates in vulval development. Genes Dev. 7, 933–947. Abstract
Mochii, M., Yoshida, S., Morita, K., Yohara, Y., and Ueno, N. (1999). Identification of transforming growth factor-b-regulated genes in Caenorhabditis elegans by differential hybridization of arrayed cDNAs. Proc. Natl. Acad. Sci. USA 96, 15020–15025. Abstract Article
Morita, K., Chow, K.L., and Ueno, N. (1999). Regulation of body length and male tail ray pattern formation of Caenorhabditis elegans by a member of TGF-β family. Development 126, 1337–1347. Abstract
Morita, K., Flemming, A.J., Sugihara, Y., Mochii, M., Suzuki, Y., Yoshida, S., Wood, W.B., Kohara, Y., Leroi, A.M., and Ueno, N. (2002). A Caenorhabditis elegans TGF-β, DBL-1, controls the expression of LON-1, a PR-related protein, that regulates polyploidization and body length. EMBO J. 21, 1063–1073. Abstract Article
Morita, K., Shimizu, M., Shibuya, H., and Ueno, N. (2001). A DAF-1-binding protein BRA-1 is a negative regulator of DAF-7 TGF-β signaling. Proc. Natl. Acad. Sci. USA 98, 6284–6288. Abstract Article
Nagamatsu, Y., and Ohshima, Y. (2004). Mechanisms for the control of body size by a G-kinase and a downstream TGFb signal pathway in Caenorhabditis elegans. Genes Cells 9, 39–47. Abstract Article
Nakano, Y., Nagamatsu, Y., and Ohshima, Y. (2004). cGMP and a germ-line signal control body size in C. elegans through cGMP-dependent protein kinase EGL-4. Genes Cells 9, 773–779. Abstract Article
Nam, S., Jin, Y.H., Li, Q.L., Lee, K.Y., Jeong, G.B., Ito, Y., Lee, J., and Bae, S.C. (2002). Expression pattern, regulation, and biological role of runt domain transcription factor, run, in Caenorhabditis elegans. Mol. Cell. Biol. 22, 547–554. Abstract Article
Newfeld, S.J., Wisotzkey, R.G., and Kumar, S. (1999). Molecular evolution of a developmental pathway: phylogenetic analyses of transforming growth factor-b family ligands, receptors and Smad signal transducers. Genetics 152, 783–795. Abstract
Nicolas, F.J., De Bosscher, K., Schmierer, B., and Hill, C.S. (2004). Analysis of Smad nucleocytoplasmic shuttling in living cells. J. Cell Sci. 117, 4113–4125. Abstract Article
Nystrom, J., Shen, Z.Z., Aili, M., Flemming, A.J., Leroi, A., and Tuck, S. (2002). Increased or decreased levels of Caenorhabditis elegans lon-3, a gene encoding a collagen, cause reciprocal changes in body length. Genetics 161, 83–97. Abstract
Oldham, S., and Hafen, E. (2003). Insulin/IGF and target of rapamycin signaling: a TOR de force in growth control. Trends Cell Biol. 13, 79–85. Article
Patterson, G.I., Koweek, A., Wong, A., Liu, Y., and Ruvkun, G. (1997). The DAF-3 Smad protein antagonizes TGF-β-related receptor signaling in the Caenorhabditis elegans dauer pathway. Genes Dev. 11, 2679–2690. Abstract
Plenefisch, J.D., DeLong, L., and Meyer, B.J. (1989). Genes that implement the hermaphrodite mode of dosage compensation in Caenorhabditis elegans. Genetics 121, 57–76. Abstract
Ren, P., Lim, C.S., Johnsen, R., Albert, P.S., Pilgrim, D., and Riddle, D.L. (1996). Control of C. elegans larval development by neuronal expression of a TGF-β homolog. Science 274, 1389–1391. Abstract Article
Riddle, D.L., Swanson, M.M., and Albert, P.S. (1981). Interacting genes in nematode dauer larva formation. Nature 290, 668–671. Abstract Article
Savage, C., Das, P., Finelli, A.L., Townsend, S.R., Sun, C.Y., Baird, S.E., and Padgett, R.W. (1996). Caenorhabditis elegans genes sma-2, sma-3, and sma-4 define a conserved family of transforming growth factorβpathway components. Proc. Natl. Acad. Sci. USA 93, 790–794. Abstract Article
Savage-Dunn, C., Maduzia, L.L., Zimmerman, C.M., Roberts, A.F. Cohen, S., Tokarz, R., and Padgett, R.W. (2003). Genetic screen for small body size mutants in C. elegans reveals many TGFb pathway components. Genesis 35, 239–247. Abstract Article
Savage-Dunn, C., Tokarz, R., Wang, H., Cohen, S., Giannikas, C., and Padgett, R.W. (2000). SMA-3 smad has specific and critical functions in DBL-1/SMA-6 TGFb-related signaling. Dev. Biol. 223, 70–76. Abstract Article
Schackwitz, W.S., Inoue, T., and Thomas, J.H. (1996). Chemosensory neurons function in parallel to mediate a pheromone response in C. elegans. Neuron 17, 719–728. Abstract Article
Schmelzle, T., and Hall, M.N. (2000). TOR, a central controller of cell growth. Cell 103, 253–262. Abstract Article
Snow, M.I., and Larsen, P.L. (2000). Structure and expression of daf-12: a nuclear hormone receptor with three isoforms that are involved in development and aging in Caenorhabditis elegans. Biochim. Biophys. Acta 1494, 104–116. Abstract
Stoyanov, C.N., Fleischmann, M., Suzuki, Y., Tapparel, N., Gautron, F., Streit, A., Wood, W.B., and Muller, F. (2003). Expression of the C. elegans labial orthologue ceh-13 during male tail morphogenesis. Dev. Biol. 259, 137–149. Abstract Article
Sulston, J.E., Albertson, D.G., and Thomson, J.N. (1980). The Caenorhabditis elegans male: postembryonic development of nongonadal structures. Dev. Biol. 78, 542–576. Abstract Article
Suzuki, Y., Morris, G.A., Han, M., and Wood, W.B. (2002). A cuticle collagen encoded by the lon-3 gene may be a target of TGF-β signaling in determining Caenorhabditis elegans body shape. Genetics 162, 1631–1639. Abstract
Suzuki, Y., Yandell, M.D., Roy, P.J., Krishna, S., Savage-Dunn, C., Ross, R.M., Padgett, R.W., and Wood, W.B. (1999). A BMP homolog acts as a dose-dependent regulator of body size and male tail patterning in Caenorhabditis elegans. Development 126, 241–250. Abstract
ten Dijke, P., and Hill, C.S. (2004). New insights into TGF-β-Smad signalling. Trends Biochem. Sci. 29, 265–273. Abstract Article
Tewari, M., Hu, P.J., Ahn, J.S., Ayivi-Guedehoussou, N., Vidalain, P.O., Li, S., Milstein, S., Armstrong, C.M., Boxem, M., Butler, M.D., et al. (2004). Systematic interactome mapping and genetic perturbation analysis of a C. elegans TGF-β signaling network. Mol. Cell 13, 469–482. Abstract Article
Thomas, J.H., Birnby, D.A., and Vowels, J.J. (1993). Evidence for parallel processing of sensory information controlling dauer formation in Caenorhabditis elegans. Genetics 134, 1105–1117. Abstract
Trent, C., Tsuing, N., and Horvitz, H.R. (1983). Egg-laying defective mutants of the nematode Caenorhabditis elegans. Genetics 104, 619–647. Abstract
Wang, J., Tokarz, R., and Savage-Dunn, C. (2002). The expression of TGFb signal transducers in the hypodermis regulates body size in C. elegans. Development 129, 4989–4998. Abstract
Xiao, Z., Watson, N., Rodriguez, C., and Lodish, H.F. (2001). Nucleocytoplasmic shuttling of Smad1 conferred by its nuclear localization and nuclear export signals. J. Biol. Chem. 276, 39404–39410. Abstract Article
Xu, L., Kang, Y., Col, S., and Massague, J. (2002). Smad2 nucleocytoplasmic shuttling by nucleoporins CAN/Nup214 and Nup153 feeds TGFb signaling complexes in the cytoplasm and nucleus. Mol. Cell 10, 271–282. Abstract Article
*Edited by Iva Greenwald. Last revised January 26, 2005. Published September 9, 2005. This chapter should be cited as: Savage-Dunn, C. TGF-β signaling (September 9, 2005), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.22.1, http://www.wormbook.org.
Copyright: © 2005 Cathy Savage-Dunn. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
§To whom correspondence should be addressed. E-mail: csavage@qc1.qc.edu
 All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.
All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.