Embryo series courtesy of Einhard Schierenberg
Embryo series courtesy of Einhard SchierenbergTable of Contents
Abstract
The C. elegans hermaphrodite vulva is an established model system to study mechanisms of cell fate specification and tissue morphogenesis. The adult vulva is a tubular shaped organ composed of seven concentric toroids that arise from selective fusion between differentiated vulval progeny. The dorsal end of the vulval tubule is connected to the uterus via a multinucleate syncytium utse (uterine-seam) cell. The vulval tubule and utse are formed as a result of changes in morphogenetic processes such as cell polarity, adhesion, and invagination. A number of genes controlling these processes are conserved all the way up to human and function in similar developmental contexts. This makes it possible to extend the findings to other metazoan systems. Gene expression studies in the vulval and uterine cells have revealed the presence of regulatory networks specifying distinct cell fates. Thus, these two cell types serve as a good system to understand how gene networks confer unique cell identities both experimentally and computationally. This chapter focuses on morphogenetic processes during the formation of the vulva and its connection to uterus.
The Caenorhabditis elegans hermaphrodite vulva is an epidermal-derived tube that connects the uterus to the external environment. Development of the vulva involves a broad range of cell biological processes that are shared among all eukaryotes. The model has been adopted for the study of signal transduction and signal integration and the subsequent transcriptional networks that direct cell differentiation. Vulva development is also a successful model for studying tissue remodeling during organ morphogenesis, including common morphogenetic processes such as invagination, lumen formation and cell fusion. The vulva provides a powerful paradigm because it has simple anatomy and rapid development and because it is easily manipulated via genetics and transgenics. Investigations with this system have revealed the function of a large number of genes and their network of interactions, including important transcription factors such as LIN-11 (LIM domain family), EGL-38 (Pax2/5/8 family of proto-oncogene), LIN-29 (zinc finger family), and COG-1 (Nkx6.1/6.2 family) (see Ririe et al., 2008 and references therein). These findings demonstrate that the vulva is a powerful system to identify and study the function of conserved genes in important cellular processes, allowing us to formulate a coherent picture of formation of a single organ.
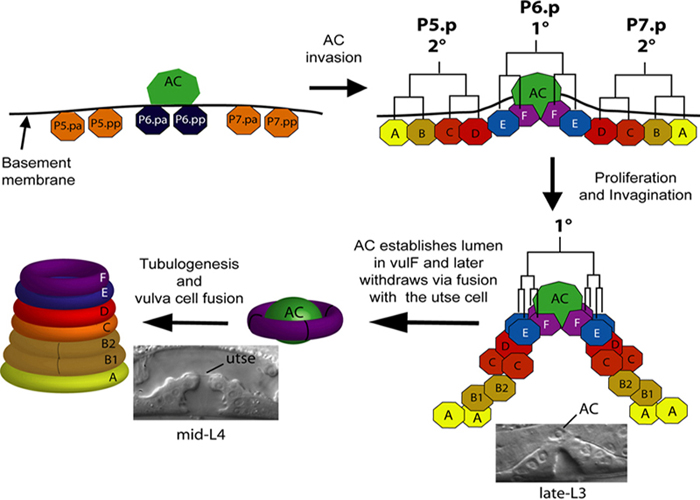
During vulval development a subset of twelve Pn.p cells acquire competence and become vulval precursor cells (VPCs). Three VPCs adopt 1° and 2° fates in response to conserved signaling pathways mediated by Ras, Wnt and Notch and divide to generate 22 progeny (Sternberg, 2005). The vulval progeny differentiate and undergo morphogenetic changes to form the adult vulva, composed of a stack of toroid-shaped cells surrounding a central lumen (Figure 1) (Sharma-Kishore et al., 1999). A specialized cell from the gonad, the anchor cell (AC), plays multiple roles in vulval development. The AC first induces VPCs to adopt 1° (P6.p) or 2° (P5.p and P7.p) fates (Sternberg, 2005). Subsequently, it controls differentiation of 1° lineage cells. The AC also patterns nearby uterine cells, via production of the DSL ligand LAG-2, which acts on LIN-12 to generate the uterine seam (utse) cell and the four uv1 cells that form the tight connection with the vulva (Newman et al., 2000; Newman et al., 1995). A connection between the uterine and vulval lumens is also formed by the anchor cell, which invades between the 1° lineage cells in a process analogous to invasive behavior of metastatic tumor cells (Sherwood and Sternberg, 2003). During this invasion process, the basement membranes between the gonad and body wall are degraded. Thus the AC patterns two neighboring tissues and ensures that the vulva and uterus connect with each other to generate a functional egg-laying system. In this chapter we summarize cellular events in the vulva and uterus that occur following VPC induction. We also present the current understanding of the roles of genes and signaling pathways that mediate the underlying processes.
|
Figure 1. Vulval cells and toroids. The P5.p, P6.p and P7.p VPCs divide three times to produce 22 progeny. The vulval progeny differentiate to give rise to 7 different cell types (vulA to vulF). The homologous cells from P5.p and P7.p lineages fuse selectively to form 7 concentric toroids that are interconnected via AJM-1-containing apical junctions. During the mid-to-late-L3 stage the AC initiates invasion into the vulval epithelium. At the early-L4 stage it can be seen establishing the lumen in vulF thereby promoting a connection between vulva and uterine lumens. By the mid-L4 stage AC has fused with π progeny to form the utse cell and its nucleus has migrated (either anteriorly or posteriorly) away from the connection between the two lumens.
The seven vulval cell types (vulA, vulB1, vulB2, vulC, vulD, vulE and vulF) are formed by the fusion of defined vulval progeny. While vulA to vulD are generated by the 2° lineage VPCs P5.p and P7.p in mirror symmetric polar patterns (ABCD and DCBA), the remaining cell types (vulE and vulF) are the progeny of the 1° lineage VPC P6.p (Figure 1). Each of these cell types has a distinct pattern of gene expression (Table 1, Figure 2) and a specialized function. For example, the vulval muscles vm1 insert between the vulE and vulF cells, while the vm2 muscles insert between the vulC and vulD cells. Additionally, the vulF cell forms the main contact with the anchor cell and induces the uv1 fate in the ventral uterine π lineage (Chang et al., 1999; Newman et al., 1996; Sherwood and Sternberg, 2003).
|
Figure 2. Gene expression patterns in vulval cells. Overview of the expression profile of 11 genes in vulval cells in wild type and mutant animals as described in literature. Expression of each gene is plotted in a matrix form where X-axis represents the vulval cell type and Y-axis represents the stage of the animal.
Table 1. Genes expressed in vulval cells between late-L3 and L4 stage (transcription factors are in bold).
| Gene | Gene family/Ortholog | Expression domain# | Key references |
|---|---|---|---|
| B0034.1 | unknown | E, F | (Inoue et al., 2002) |
| bam-2 | Neurexin-related | F | (Colavita and Tessier-Lavigne, 2003) |
| C55C3.5 | unknown | F | (Inoue et al., 2005) |
| cdh-3 | FAT-like cadherin | C, D, E, F | (Inoue et al., 2005; Pettitt et al., 1996) |
| ceh-2 | HOX | B1, B2, C | (Inoue et al., 2002) |
| cog-1 | Nkx6.1/6.2 | C, D, E, F, and occasionally in A and B | (Palmer et al., 2002) |
| daf-6 | Patched-related | E, F | (Perens and Shaham, 2005) |
| dhs-31 | Predicted short chain-type dehydrogenase | B1, B2, D | (Inoue et al., 2002) |
| egl-17 | Fibroblast growth factor-like | C, D | (Burdine et al., 1998) |
| egl-26 | Acyltransferase-like | B, D, E | (Hanna-Rose and Han, 2002) |
| egl-38 | Paird-box domain (Pax2/5/8) | F | (Rajakumar and Chamberlin, 2007) |
| grd-5 | Groundhog hedgehog | B1, B2, D | (Hao et al., 2006) |
| grd-12 | Groundhog hedgehog | C | (Hao et al., 2006) |
| grl-4 | Ground-like hedgehog | A, B1, B2, D | (Hao et al., 2006) |
| grl-10 | Ground-like hedgehog | A, B1, B2 | (Hao et al., 2006) |
| grl-15 | Ground-like hedgehog | B1, B2, C, D, E | (Hao et al., 2006) |
| grl-25 | Ground-like hedgehog | A | (Hao et al., 2006) |
| grl-31 | Ground-like hedgehog | F | (Hao et al., 2006) |
| lin-3 | Epidermal growth factor-like | F | (Chang et al., 1999) |
| lin-11 | LIM-HOX | A, B1, B2, C, D | (Gupta and Sternberg, 2002) |
| lin-29 | C2H2-type Zn finger | All cells | (Bettinger et al., 1997) |
| lin-39 | HOX | A | (Wagmaister et al., 2006) |
| nas-37 | Metalloprotease | B | (Ririe et al., 2008) |
| nhr-67 | Zn finger NHR/Tailless | A, B, C | (Fernandes and Sternberg, 2007) |
| nhr-113 | Zn finger NHR | A | (Ririe et al., 2008) |
| npax-2 | Paird-box domain | C, D | (Reece-Hoyes et al., 2007), (Ian Hope, pers. comm.) |
| pax-2 | Paird-box domain (Pax2/5/8) | D | (Fernandes and Sternberg, 2007) |
| pepm-1 | Peptidase | C, D, E, F | (Inoue et al., 2002) |
| sqv-4 | UDP-glucose dehydrogenase | C, D, E, F | (Hwang and Horvitz, 2002) |
| syg-2 | Cell adhesion Ig domain | E, F | (Shen et al., 2004) |
| unc-53 | Nuclear pore membrane and/or filament interacting like (POMFIL) | C | (Stringham et al., 2002) |
| vrk-1 | Protein kinase (vaccinia-related) | All cells | (Klerkx et al., 2009) |
| zmp-1 | Zn metalloproteinase | A, D, E | (Inoue et al., 2002) |
|
#refers to expression in vulA to vulF vulval cell types (in short, A to F). In some cases expression in B1 and B2 was not clearly identified. These are shown as B. |
|||
The 1° lineage generates two cell types, vulE and vulF, in a mirror symmetric pattern EFFE around the anchor cell (Figures 1 and 3). The anchor cell is necessary for establishing this pattern after it induces the 1° VPC via the LET-23 (Epidermal Growth Factor Receptor, EGFR)-LET-60 (RAS)-mediated pathway (Wang and Sternberg, 2000) (Figure 3). WNT receptor LIN-17 plays a role as well, but the phenotype of a lin-17 mutation has only been seen in the absence of the anchor cell (Wang and Sternberg, 2000). The relative timing of the signaling involved in these events has not been resolved. The identities of vulE and vulF are specified by the expression of an array of genes including egl-26 (acyltransferase-like), egl-38 (PAX2/5/8 family), and cog-1 (Nkx3.1 family) (Estes et al., 2007; Hanna-Rose and Han, 2002; Inoue et al., 2005; Palmer et al., 2002; Rajakumar and Chamberlin, 2007). Although the precise roles of these genes are not fully resolved, both cell autonomous and non-autonomous mechanisms are likely to play roles.
|
Figure 3. Patterning of the 1° lineage. A. Intact, wild-type 1° lineage has two F cells near the anchor cell and two more distal E cells. B. After AC ablation, the patterns are variable. C. In a lin-12(lf) mutant in which all ventral uterine cells but one anchor cell have been ablated, and in which P7.p has been ablated to keep only two 1° lineages, there is a normal pattern at the anchor cells, but a variable distal pattern. D. Model posits an AC-mediated signal via RAS to specify vulF cells near the AC. In addition, vulE and vulF signal each other to ensure that correct fates are specified. Data and model summarized previously (Wang and Sternberg, 2000).
Each of the 2° lineages (P5.p and P7.p) generates seven progeny that produce five vulval cell types, vulA to vulD (see above). The specification of the 2° fate is dependent on the LIN-12-mediated lateral signal (Greenwald, 2005). A MAP kinase phosphatase LIP-1 that acts downstream of LIN-12, antagonizes MAP kinase MPK-1 activity in P5.p and P7.p, allowing them to adopt a 2° fate (Berset et al., 2001). The negative feedback on the kinase cascade downstream of the LET-60 (Ras) - LIN-45 (Raf) signaling module also allows LET-60 to specifically signal via RGL-1 (Ral GEF) and RAL-1 to promote 2° fate in P5.p and P7.p (Zand et al., 2011). Wnt signaling (involving LIN-17/Frizzled and PRY-1/Axin) is also involved in 2° fate determination (Eisenmann, 2005; Seetharaman et al., 2010), in part by interacting with LIN-12 signaling (Seetharaman et al., 2010). In addition to fate specification, Wnt signaling also plays a role in reversing the polarity of P7.p such that a symmetrical vulval invagination can be formed (see below). A Ryk family tyrosine kinase-related protein LIN-18 functions as a co-receptor of LIN-17 in this process (Inoue et al., 2004).
Many genes have been identified that act downstream of the signaling pathways (Ras, Notch, and Wnt) to regulate identities of specific vulval cell types. These include conserved genes encoding transcription factors and other proteins (Table 1). Genetic epistasis and transgene expression experiments have revealed a functional hierarchy in a few cases but the overall transcriptional network underlying vulval differentiation appears to be complex (Figure 2). This is in part due to seven different cell types and their unique gene expression programs (Figure 2). Cis regulatory analysis has also revealed a similar extent of complexity in the specification of the vulval cell types (Cui and Han, 2003; Kirouac and Sternberg, 2003 ; Marri and Gupta, 2009; Ririe et al., 2008). Below we discuss some of the genes that have been studied in some detail and shown to play major roles in vulval development.
The LIM Hox transcription factor lin-11 is a key regulator of vulval cell fates (Ferguson et al., 1987; Gupta et al., 2003). It is expressed in both the 1° fate (during mid-L3) and 2° fate lineages (during L4) and promotes fates in a cell autonomous manner (Gupta et al., 2003). During the L4 stage lin-11 controls differentiation and morphogenesis of all P5.p and P7.p progeny, as judged by expression of markers such as zmp-1, cdh-3 and egl-17 (Gupta et al., 2003). This is mediated in part by a lin-11 expression level that is higher in vulC and vulD compared to vulA, vulB1 and vulB2 cells (Freyd, 1991 ; Gupta et al., 2003). Promoter dissection experiments have revealed the presence of Ras, Notch and Wnt response elements in the lin-11 5’ UTR suggesting that lin-11 is regulated by all three signaling pathways (Gupta and Sternberg, 2002).
Another transcription factor lin-29 (Zn finger family) is also expressed in 1° and 2° lineage vulval cells and controls cell fate specification (Bettinger et al., 1997). lin-29 mutants show reduced lin-11::GFP fluorescence in 2° vulval cells suggesting that lin-29 positively regulates lin-11 expression (Ririe et al., 2008). However the nature of the interactions between the two genes is complex because they affect target genes differently. As a regulator of developmental timing lin-29 was earlier thought to act within a temporal window (Inoue et al., 2005; Newman et al., 2000) but this hypothesis has been weakened by analysis of additional targets (Ririe et al., 2008). Specifically, the mid-L4 expression of lin-3 in vulF cells is not dependent upon LIN-29 while dhs-31, expressed later in the adult, does require LIN-29 function (Ririe et al., 2008). lin-29 is also involved in the development of the uterus and the seam cells and their interconnections (Newman et al., 2000).
Besides lin-11 and lin-29, several other genes have been identified that function in subsets of vulval progeny. egl-26 encodes an acyltransferase-like protein that is expressed in the membranes of vulE cells and is proposed to promote vulF morphogenesis via an unknown cell-cell interaction mechanism (Hanna-Rose and Han, 2002). EGL-26 is located in the apical membrane region of cells and this localization is necessary for proper function of the protein. Mutations in egl-26 do not cause defects in vulE toroid formation as judged by the cell junction marker ajm-1. Instead they specifically affect vulF morphology (Estes et al., 2007).
Other vulF specificity determinants include the PAX2/5/8-like transcription factor EGL-38 and the EGF family ligand LIN-3 both of which are specifically localized to vulF. Mutations in egl-38 cause a failure of vulF cells to separate leading to a physical block in the egg-laying passage (Chamberlin et al., 1997; Chang et al., 1999; Rajakumar and Chamberlin, 2007). egl-38 expression is positively autoregulated (Rajakumar and Chamberlin, 2007). egl-38 promotes expression of lin-3 and represses cog-1 in these same cells (Chang et al., 1999; Inoue et al., 2005). In addition, the vulE marker zmp-1 is ectopically expressed in vulF in some of the egl-38 animals, possibly due to an F to E fate transformation (Inoue et al., 2005). However the pattern of another vulF marker bam-2 remains unchanged suggesting that egl-38 does not cause a widespread change in the vulF transcriptional program (Rajakumar and Chamberlin, 2007).
The vulF expression of egl-38 and lin-3 are promoted by the Zn finger/nuclear hormone receptor (NHR) family member nhr-67 (Ririe et al., 2008). In addition nhr-67 also regulates several other targets in vulval cells. It promotes zmp-1 (in vulA) and inhibits egl-17 (in vulF), cog-1 and ceh-2 (both in vulE and vulF) (Fernandes and Sternberg, 2007). The relationship between cog-1 and nhr-67 appears to be complex. The two genes negatively autoregulate each other's expression in vulE and vulF cells. Interestingly, the early expression of nhr-67 in vulval cells is negatively autoregulated since it can be seen only in the absence of nhr-67 function (Fernandes and Sternberg, 2007). Thus, genes other than zmp-1 are likely to be indirect targets of nhr-67. The complex genetic interactions of nhr-67 and other vulval cell type regulators and targets are more easily rationalized with nhr-67 acting in the vulval lineages, but its strong expression in the anchor cell suggests that it acts in the anchor cell to specify vulval cell fates. Definitive site-of-action experiments are necessary to clear up this key issue.
The Nkx6.1 transcription factor family member cog-1 is dynamically expressed in vulval cells. It is first observed in vulE and vulF precursors during the L3 stage. At a later stage (L4) expression is detected in 2° lineage cells in a pattern similar to lin-11. Thus, vulC and vulD precursors show a higher level of expression compared to others. The cog-1 locus encodes two transcripts that are thought to function differently in vulval cell fate specification. This is based on the phenotypic analysis of cog-1 mutations that affect only one (sy607) or both (sy275) transcripts (Palmer et al., 2002). The two alleles differentially affect vulval cell fates as judged by cell lineage and marker gene (ceh-2 and egl-17) expression studies (Inoue et al., 2005; Palmer et al., 2002). The cog-1(sy275) animals show a physical block in the vulval-uterine connection, which is caused by a failure in the separation of vulF cells (Palmer et al., 2002). This phenotype is similar to egl-26 and egl-38, but the relationship among cog-1, egl-26 and egl-38 is unknown.
Apart from the above genes a few others, such as lin-26 (C2H2 Zn finger) (Eisenmann and Kim, 2000) and bed-3 (BED type Zn finger) (Inoue and Sternberg, 2010), also control vulval cell fates but their roles have not been studied in much detail.
The patterned vulval cells ultimately form an epithelial tube with a central lumen that allows passage of eggs. Tubulogenesis begins even before all cell divisions are complete and is not completed until the mid-to late L4 stage, hours after the last cell divisions. During morphogenesis the vulval cells orient and migrate towards the anchor cell, undergo epithelial invagination, and fuse to form seven toroids (see below). After the vulval tube forms, it everts during the L4 molt to form the mature vulva, required for regulated egg laying and mating with males.
The epithelial vulval cells are highly polarized with their apical edges adjacent to the lumen. While there is no evidence that the par genes, which are required for polarizing cells in the embryo and elsewhere, are involved in setting vulval cell polarity, experiments to examine their function during vulval development are complicated by maternal effects and early larval lethality of null alleles. Thus, it remains to be seen if par genes function during polarity establishment in the vulva. One of the par family members, par-1, was earlier found to be expressed in vulval cells and is necessary for morphogenesis (Hurd and Kemphues, 2003), however its cellular role in vulva formation remains unknown.
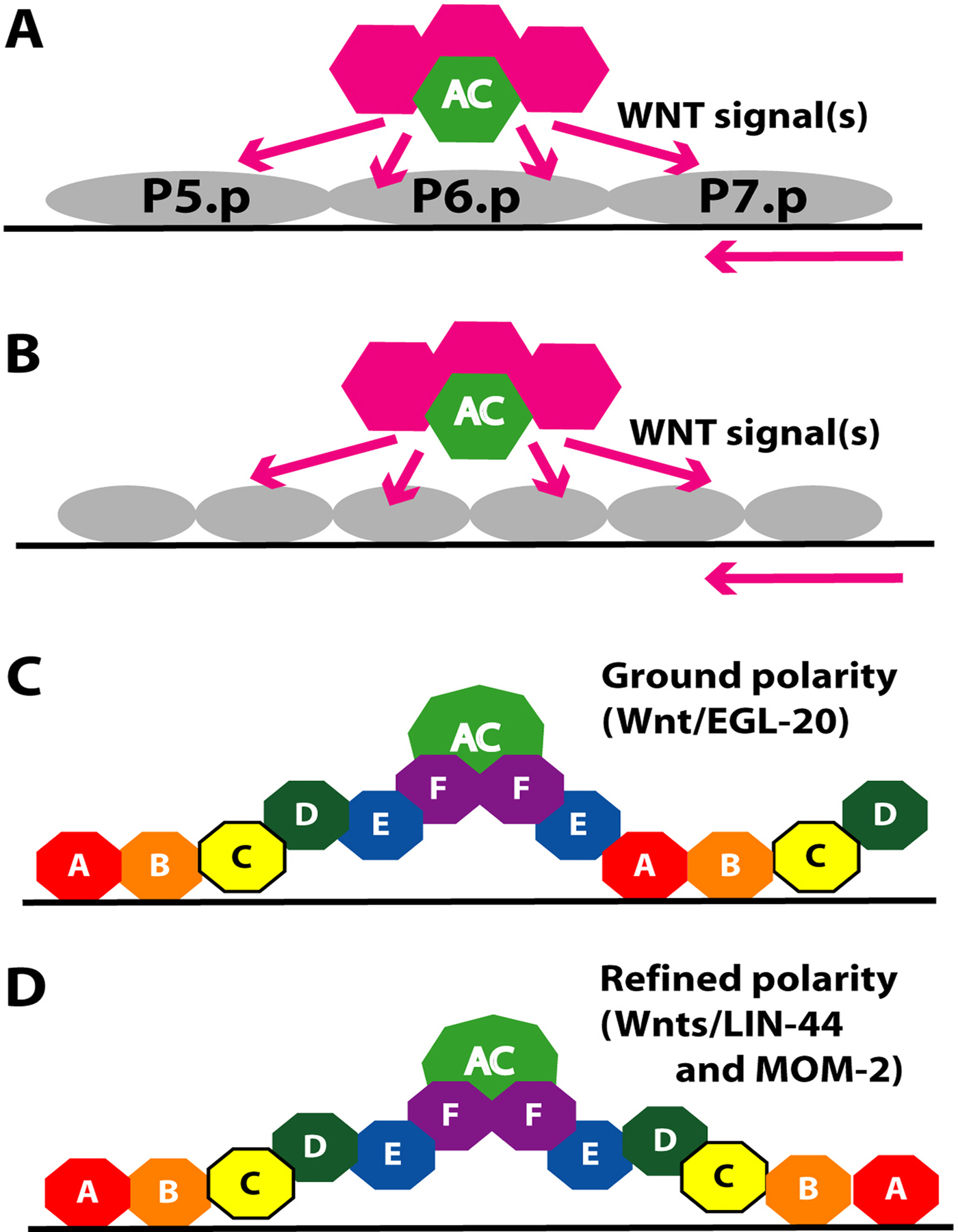
Wnt signaling is another major regulator of cell polarity decisions in C. elegans. In the vulva Wnt signaling controls the orientation of 2° lineage cells (Figure 4). A default “ground state” polarity is initially established by Wnt/EGL-20 in which P5.p and P7.p progeny orient towards the posterior. Subsequently another Wnt signal from the anchor cell reverses the default polarity of the P7.p lineage and reinforces the polarity of the P5.p lineage (Deshpande et al., 2005; Green et al., 2008; Inoue et al., 2004). As a result, the cell division pattern and cell fates of the P5.p and P7.p lineages are mirror images, ABCD and DCBA respectively (Figure 4). The asymmetry within each of these lineages is independent of the gonad, as for example, a single VPC in a gonad-ablated let-23(gf) mutant has asymmetry (Katz et al., 1996). Three WNT signaling pathways act together to specify polarity (Green et al., 2008; Inoue et al., 2004). (1) The WNT protein EGL-20, likely due to its expression in the tail, acts as a long-range signal and orients P5.p and P7.p to the posterior (Green et al., 2008). This pathway requires WNT receptor CAM-1 and membrane protein VANG-1, although the detailed mechanism is not known. (2) WNT protein MOM-2, which is expressed in the anchor cell, helps to reverse P7.p polarity and orient P5.p and P7.p towards the anchor cell. MOM-2 acts via the Ryk-type receptor LIN-18. (3) WNT protein LIN-44, which is also expressed in the anchor cell, cooperates with MOM-2, but acts via the Frizzled-type receptor LIN-17 (Ferguson et al., 1987; Inoue et al., 2004; Sawa et al., 1996; Sternberg and Horvitz, 1988). This signal antagonizes the action of EGL-20 for P7.p, and reinforces the action of EGL-20 for P5.p. In addition, lin-44 transcript is also observed in the anchor cell region based on in situ hybridization (Y. Kohara, personal communication).
|
Figure 4. Polarity of P7.p 2° lineage. Multiple WNT signals from the tail region (not shown), anchor cell (green) and other gonadal cells (pink) help establish the correct polarity of the P5.p and P7.p lineage cells. Wnt/EGL-20 originating from the tail region orients cells towards the posterior (ground polarity). This pattern is reversed in the P7.p lineage by Wnts/LIN-44 and MOM-2 (refined polarity) such that it has an DCBA pattern rather than the default ABCD pattern as does P5.p. A. early-L3. B. mid-L3. C and D. late-L3. Model is based on published studies (Deshpande et al., 2005; Green et al., 2008; Inoue et al., 2004).
The levels of nuclear POP-1 (TCF/LEF family), a WNT target, correlate with WNT signaling: a High-Low (HL) pattern is observed in P5.p daughters and HLHL pattern in P5.p granddaughters, while a LH then a LHLH pattern is seen in the case of P7.p (Deshpande et al., 2005). Thus, the pattern of POP-1 protein localization in the P7.p lineage is reversed from the default by WNT signaling through LIN-17 and LIN-18. This ultimately leads to reversal of P7.p polarity and giving rise to a vulva with single invagination where the two halves are mirror symmetric. A reporter for POP-1 activation of transcription, POPTOP (Green et al., 2008), indicates that P7.p polarity reversal is largely independent of POP-1-mediated transcriptional activity. Reduction of function in the SP1 family protein SPTF-3 causes a similar P7.p polarity reversal, leading to the hypothesis that SPTF-3 interacts with Wnt signaling in some way. However, this remains to be investigated (Ulm et al., 2011).
At the end of L3 even before vulval cell divisions are complete, the distal vulval cells begin to move, migrating under their proximal neighbors and pushing the proximal cells dorsally. Invagination of these epithelial cells continues during the early L4 larval stage, forming a rudimentary vulva lumen beneath the anchor cell and between the anterior and posterior cells of the vulva. These morphogenetic movements involve cytoskeletal reorganization.
At least two different G protein-mediated signaling pathways facilitate morphogenetic changes in the vulva. A member of the ARL2 family of small GTPases called EVL-20 controls invagination of vulval cells (Antoshechkin and Han, 2002). evl-20 mutants exhibit defects in microtubule reorganization suggesting that vulval abnormalities are caused by a failure to regulate cytoskeletal dynamics. Two other GTPases of the Rac family, encoded by mig-2 and ced-10, are also necessary for appropriate division and migration of vulval cells (Kishore and Sundaram, 2002). Their activities are likely to be regulated by the Trio-like guanine nucleotide exchange factor UNC-73.
Semaphorin SMP-1 and its Plexin receptor PLX-1 represent yet another signaling pathway that acts downstream of Ras signaling and in parallel to Rac signaling to control vulval cell movements and invaginations (Dalpe et al., 2005; Pellegrino et al., 2011). The expression of SMP-1 in the VulD, E and F cells is regulated by VAB-23 in response to the inductive signal from the AC. The inductive signal results in activation of the homeobox protein LIN-39, which binds directly to the vab-23 promoter. VAB-23, in turn, binds the smp-1 promoter and promotes SMP-1 expression (Pellegrino et al., 2011). Based on mutant phenotypes and expression data it has been proposed that PLX-1 and SMP-1 mediate orderly attraction of vulval cells (starting from vulF) to form toroids.
The invagination of vulval cells results in a space (lumen) (Figure 1) that expands during L3 and L4 and is filled with fluid. Lumen expansion is mediated by proteoglycans. So far eight sqv genes (sqv-1 to sqv-8) have been identified that regulate expansion of the vulval lumen (Herman et al., 1999). Hermaphrodites with decreased sqv function develop only a rudimentary lumen (SQuashed Vulva phenotype). The sqv genes encode a nucleotide-sugar transporter (sqv-7) (Berninsone et al., 2001; Herman and Horvitz, 1999) and glycosaminoglycan biosynthetic enzymes that are required for synthesis of chondroitin and heparan (Herman and Horvitz, 1999; Hwang and Horvitz, 2002; Hwang and Horvitz, 2002; Hwang et al., 2003; Hwang et al., 2003). Accumulation of hygroscopic chondroitin sulfate in the vulval lumen is proposed to draw water into the lumen, generating the pressure to hold the lumen open during development (Hwang et al., 2003). The sqv genes are expressed broadly in the vulva with the exception of sqv-4, which is expressed in vulC, vulD, vulE and vulF (Hwang and Horvitz, 2002).
The mature vulva is composed of seven concentric toroids that form as each vulval cell extends processes laterally around the developing vulval lumen, contacting each other and subsequently fusing in a selective manner (Sharma-Kishore et al., 1999) (Figure 1). The multinucleate vulval toroids vulA, vulC, vulD, vulE and vulF form via homotypic fusion. The vulB1 and vulB2 cells persist as rings composed of two distinct cells (Sharma-Kishore et al., 1999).
Two FF (fusion failure) family genes, eff-1 and aff-1, function independently to control cell-fusion events in the vulva (Mohler et al., 2002; Sapir et al., 2007). Both encode transmembrane proteins with similar domain organization that are conserved in nematodes. eff-1 is expressed in a subset of vulval cells and is required for fusion of some of the vulval toroids but not vulA and vulD (Sapir et al., 2007). The expression of eff-1 is regulated by VAB-23 in the vulva. When VAB-23 function is reduced, eff-1 is ectopically expressed, resulting in ectopic fusions and fewer toroids (Pellegrino et al., 2011). aff-1 is expressed in the vulD precursors cells and is required for fusion of the vulD cells and possibly the vulA cells (Sapir et al., 2007). aff-1, but not eff-1, is also involved in the AC-utse cell fusion event (see 4.5). Given that AFF-1 has no apparent role in the primary vulval lineage and that both fusogens mediate homotypic fusion events, it is difficult to envision how eff-1 alone mediates the vulE to vulE and vulF to vulF specific fusions within the primary lineage. The fusogens may act combinatorially or other factors may regulate their activities. Alternatively strict control in the timing of expression between vulE and vulF could promote specific fusions within the primary lineage.
In addition to the FF genes, components of the vacuolar ATPase complex, fus-1, vha-1 and vha-12 (e, c and B subunits, respectively), have also been found to control cell fusion in the embryo by repressing eff-1 activity (Kontani et al., 2005). However their roles in the vulva have not been examined.
The vulval epithelium organizes the development of the muscles and neurons that mediate egg laying (Li and Chalfie, 1990). This is most obviously seen in dig-1 mutants in which the gonad is shifted anteriorly and the vulva, vulval muscles and neurons are all similarly shifted anteriorly (Thomas et al., 1990). Specific gene products expressed in subsets of vulval cells direct details of development of individual neurons. bam-2 is necessary for axonal branch termination of the VC motoneurons, and is expressed in the vulF cells (Colavita and Tessier-Lavigne, 2003). syg-1 and syg-2 are required for synapse formation by the HSN neurons (Shen and Bargmann, 2003; Shen et al., 2004). syg-2 is expressed in vulE cells and syg-1 in the HSNs.
The anchor cell is the key organizer of the vulva, the ventral uterus and the vulval-uterine connection (Figures 1 and 5). Once specified during the L2 molt (see WormBook chapter Hermaphrodite cell-fate specification), the anchor cell induces and patterns the vulva. The anchor cell also patterns the ventral uterus, inducing the nearest six of twelve ventral uterine cell (VU) granddaughters to adopt a π fate via production of the LAG-2 signal (Figure 5). Two cell types, the syncytial uterine seam (utse) cell and the four uterine vulva 1 (uv1) cells, are derived from the π lineage. In addition to its role as a signaling center, the anchor cell also plays important roles during morphogenesis of the connection between the vulva and the uterus (see 4.4 and 4.5).
|
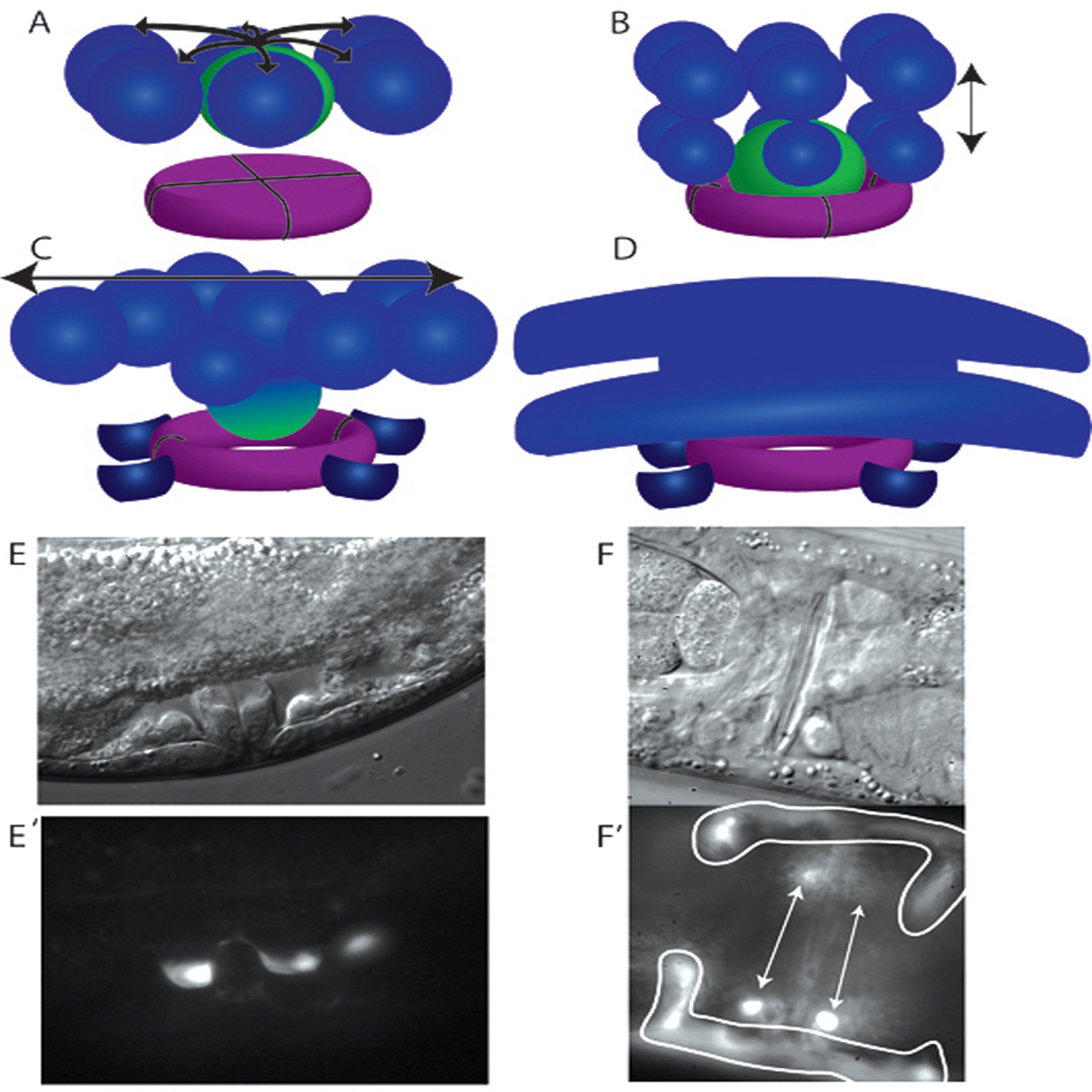
Figure 5. Uterine π cell developmental progression. (A) The six ventral uterine cells (blue) surrounding the AC (green) adopt a π fate in response to signaling (arrows) from the anchor cell. (B) The π cells divide once with a dorsal-ventral orientation (double headed arrow), producing twelve π cells. The AC invades the vulF precursors, forming a lumen in vulF (purple). (C) The ventral, outer π progeny change shape and adopt the uv1 fate (dark blue). The uv1 cells remain in close proximity to vulF. The remaining eight π progeny fuse with each other and the retreating AC to form the uterine seam (utse) cell, which expands in the anterior posterior plane (double headed arrow) to form the mature utse cell. (D) The mature utse cell has a thin cytoplasm that overlies the vulva and the uv1 cells and arms that extend laterally to the anterior and the posterior along the lateral wall of the animal. The arms curve ventrally to flank the vulva. The nine nuclei of the utse cell are found at the ends of the arms (not shown). (E and E′) The uv1 cells have adopted a sterotypical morphology with a concave dorsal surface (E′) by the time of vulval eversion (E). The uv1 cells are visualized with a Pida-1::GFP transgene, inIs179. (F and F′) Both the uv1 cells (arrows) and uste cell (outlined) are visibile in this ventral view of a young adult animal with a mature vulva. Cells are visualized with a Pexc-9::GFP transgene.
Anchor cell production of the LAG-2 signal is received in the VU cell granddaughters via the LIN-12 receptor (Newman et al., 2000; Newman et al., 1995) (Figure 5). Genetic experiments have shown that while lag-2 is regulated by the Zn-finger gene lin-29, lin-12 expression depends on the NHR family member nhr-67 (Newman et al., 2000; Verghese et al., 2011). In epistasis analysis, the Evi1 homolog egl-43 is required downstream of or in parallel to lin-12 to specify π fate (Rimann and Hajnal, 2007), and egl-43 is likely directly regulated by the lin-12 coactivator lag-1 (Hwang et al., 2007). The π cells undergo a single round of dorsal-ventral oriented cell divisions and are easily distinguished from the uninduced VU granddaughter cells, which undergo two rounds of cell division, primarily in the anterior posterior plane (Newman et al., 1996). The twelve resulting π progeny give rise to the single syncytial utse cell and the four uv1 cells, which occupy positions between the vulva and the gonad and surrounding the connection between the vulval and uterine lumens (Figure 5).
Eight of the π progeny fuse and undergo morphogenesis to form the uterine seam cell. This cell underlies the uterus and connects to the lateral seam cells, holding the uterus in place. The LIM domain transcription factor LIN-11 and the Sox domain transcription factor EGL-13 (a.k.a. COG-2) are expressed in all π cells and their daughters (Hanna-Rose and Han, 1999; Newman et al., 1999). Although π cells are still generated in either the egl-13 or the lin-11 mutant, each of these transcription factors is required for full differentiation of the utse cell (Cinar et al., 2003; Newman et al., 1999). In the absence of differentiation and/or maintenance of the utse cell fate in a lin-11 or an egl-13 mutant, the anchor cell and utse do not fuse (Hanna-Rose and Han, 1999; Newman et al., 1999) (see 4.5 AC-utse fusion). The uterine π lineage enhancer of the lin-11 gene has binding sites for FOS-1, LAG-1 and EGL-43, each of which is required for the expression of lin-11 in the π lineage cells (Marri and Gupta, 2009). Sumoylation of LIN-11 is also necessary for its function in π lineage specification (Broday et al., 2004). egl-13 is co-regulated by LAG-1 and FOS-1, which each bind its promoter (Oommen and Newman, 2007). The current model for utse differentiation suggests that activation of LIN-12 leads to expression of EGL-43 (Figure 6). Then LAG-1 and EGL-43 in conjunction with FOS-1 jointly regulate expression of lin-11 and egl-13.
|
Figure 6. Regulation of uterine π cell induction. The anchor cell expresses DSL-type ligand LAG-2 in a LIN-29-dependent manner and signals presumptive π cells via LIN-12. LIN-12 activation leads to expression of lin-11 and egl-13, two transcription factors necessary for utse development, including fusion of the anchor cell with utse cell. Model adapted from published studies (Cinar et al., 2003; Marri and Gupta, 2009; Newman et al., 1999; Newman et al., 2000; Verghese et al., 2011).
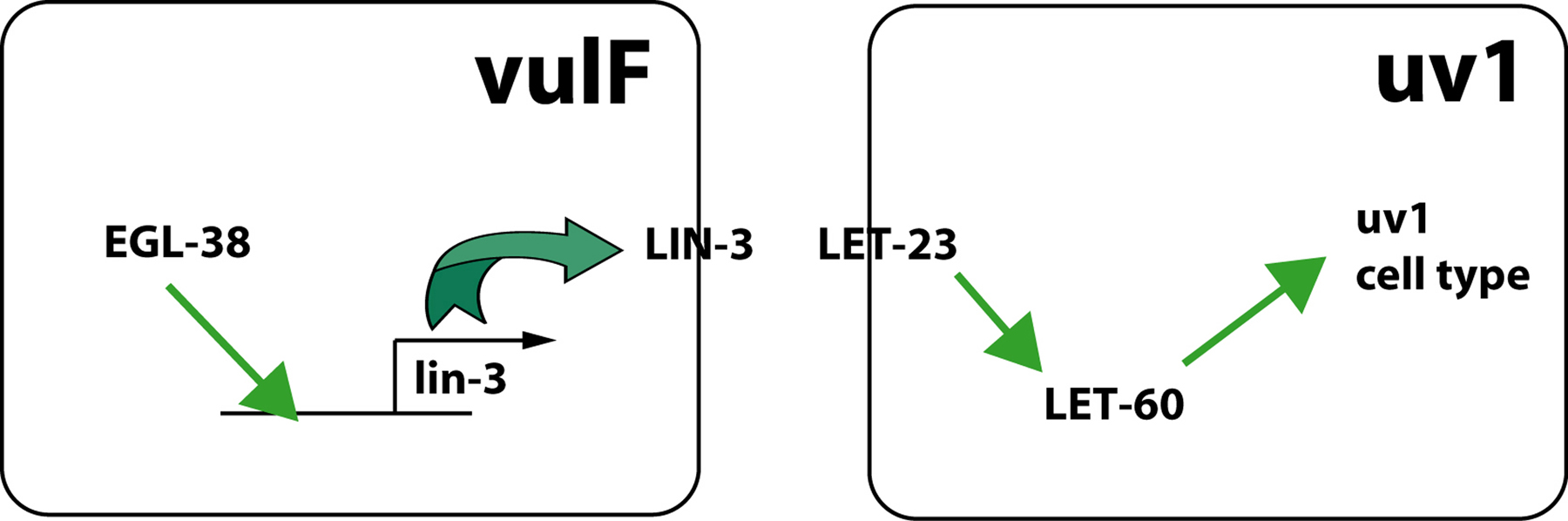
The remaining four π progeny generate the uv1 cells that lie under the uterus in proximity to the side and the top of the vulF cells (Figure 5). Specification of the uv1 cells first requires specification of the π lineage and then a second signaling event. The neighboring vulF cells express LIN-3 in an egl-38-dependent manner, and the uv1 cells respond to LIN-3 via the LET-23 receptor (Chang et al., 1999) (Figure 7). The eight π progeny that give rise to the utse do not appear to express LET-23, and a gain-of-function mutation in LET-23 does not cause a utse to uv1 fate transformation (Huang and Hanna-Rose, 2006). However, the mechanism for generating this initial difference in expression pattern between the utse and uv1 precursors is not known and the upregulation of let-23 in the uv1 cells may be in response to the lin-3 signal (Chang et al., 1999). It has been suggested that the utse fate is the default fate in the π lineage since uv1 cells adopt the utse fate in certain mutant situations (e.g., in egl-38 mutants) (Rajakumar and Chamberlin, 2007) whereas no utse to uv1 fate transformation has been observed.
|
Figure 7. uv1 specification. The vulF cell expresses LIN-3 and signals the presumptive uv1 cells via LET-23 and LET-60. egl-38 is expressed in vulF and is necessary for lin-3 expression in the same cell (Chang et al., 1999; Rajakumar and Chamberlin, 2007). Activated LET-23 bypasses this requirement for EGL-38 function.
The uv1 cells are hypothesized to be mechanosensory neurosecretory cells (Alkema et al., 2005; Jose et al., 2007; Kim and Li, 2004). They express TRP channels that negatively regulate egg laying. Dominant negative mutations in ocr-2, a TRP channel subunit, have increased rates of egg laying under high osmolarity conditions (Zhang et al., 2010) and a concommitant decrease in number of eggs held in the uterus (Jose et al., 2007). This egg-laying function requires expression of ocr-2 within the π lineage uv1 and/or utse cells (Jose et al., 2007). TRP channel subunits appear to be co-expressed only in the uv1 cells and not the utse. Nonetheless, animals engineered to have no uv1 cells (by induction of let-23 RNAi specifically in the π lineage) have no detectable increase in egg laying under high osmolarity conditions and no decrease in egg accumulation in the uterus (Johnson et al., 2009) (W. Hanna-Rose, unpublished). These animals also have no detectable defects in morphogenesis of the vulval-uterine connection (Johnson et al., 2009). Thus, the role of the uv1 cells is not clear.
In addition to inducing π cell fate, the anchor cell also promotes formation of a direct physical connection between the vulva and the uterus via other cell biological processes. The gonad and the vulval epithelium are originally separated by basement membranes secreted by each tissue. The anchor cell responds to a netrin signal from the ventral nerve cord by assembling an invasive membrane rich in actin and phospholipid PI(4,5)P2 adjacent to the basement membranes (Ziel et al., 2009). The INA-1-PAT-3 integrin heterodimer is required in the anchor cell for assembly of the invasive membrane and subsequent invasion of the basement membrane (Hagedorn et al., 2009). After the invasive membrane is formed, the anchor cell degrades and removes the basement membranes and invades, coming to lie with its basolateral side embedded in the apex of the vulva (Figure 1) (Sherwood et al., 2005; Sherwood and Sternberg, 2003). Three transcription factors, fos-1a, egl-43 and nhr-67, control the invasive behavior of the anchor cell (Hwang et al., 2007; Rimann and Hajnal, 2007; Sherwood et al., 2005; Verghese et al., 2011). fos-1a, a component of the AP-1 transcriptional regulator, is strictly required for anchor cell invasion and acts upstream of egl-43. Some downstream targets of fos-1a and egl-43 with roles in invasion include the metalloproteinase zmp-1, him-4 hemicentin and the Fat-like protocadherin cdh-3. These genes have partially penetrant invasion defects, suggesting redundant function in invasive processes.
Anchor cell invasion usually occurs only in the presence of the vulva, but the anchor cell can invade in ~20% of vulvaless animals (Sherwood and Sternberg, 2003). During invasion, the anchor cell extends processes towards the developing 1° lineage. The anchor cell apparently responds to cues from vulF (and possibly negative cues from vulE) to locate the vulF cells (Sherwood and Sternberg, 2003). When the anchor cell is not ideally situated above the vulF precursor cells, it can send its process by the vulE cells to locate the vulF cells. The as-yet-unidentified signal from the vulva to the invading anchor cell acts at a distance because a distant 1° VPC can attract extension of the anchor cell. A large-scale screen for pro-invasive genes has identified other genes that play no role in invasive membrane formation and appear to be outside the fos-1a pathway, indicating that other aspects of anchor cell invasion in addition to the vulval cue have yet to be elucidated (Matus et al., 2010).
A second function of anchor cell invasion is to contribute to lumen formation in the vulva. The anchor cell, which has breached the basement membranes, sits in a pocket surrounded by the four vulF precursors (Figure 5) (Estes and Hanna-Rose, 2009). One of the anchor cell processes that has extended towards vulF penetrates between the vulF precursors and is followed by a significant portion of the anchor cell body (Figure 1). The basolateral surface of the anchor cell appears to contact the vulval lumen (Sherwood and Sternberg, 2003). While no defects in vulF specification have been detected when the anchor cell fails to occupy this position, the lumen in vulF fails to form (Estes and Hanna-Rose, 2009). Thus, the anchor cell is proposed to be key to opening the dorsal side of the vulva lumen potentiating a connection to the uterine lumen.
After the anchor cell completes its signaling and invasion functions during morphogenesis, it sits in a position that blocks the connection between the developing uterine and vulval lumens. To clear this opening, the utse cell syncytium fuses to the anchor cell during the L4 stage, leaving a thin process of utse cell cytoplasm overlying the vulva (Figures 1 and 5) (Kimble and Hirsh, 1979; Newman et al., 1996). This hymen is ruptured when the first egg is laid, establishing a direct connection between the uterine and vulval lumens. The fusogen AFF-1 is necessary for AC-utse fusion as well as other cell fusion events in C. elegans development (Sapir et al., 2007). aff-1 is expressed in a FOS-1A-dependent manner in both the AC and the utse, promoting homotypic fusion (Sapir et al., 2007). A missense mutation in the N-ethylmaleimide-sensitive factor NSF-1 specifically abrogates anchor cell-utse cell fusion, but not other cell fusions (Choi et al., 2006), and the nsf-1L isoform of the nsf-1 gene acts in the anchor cell to promote fusion. Whereas, AFF-1 is a membrane protein with an extracellular domain that mediates homotypic fusion, the role of the intracellular vesicle trafficking protein NSF-1 is not clear.
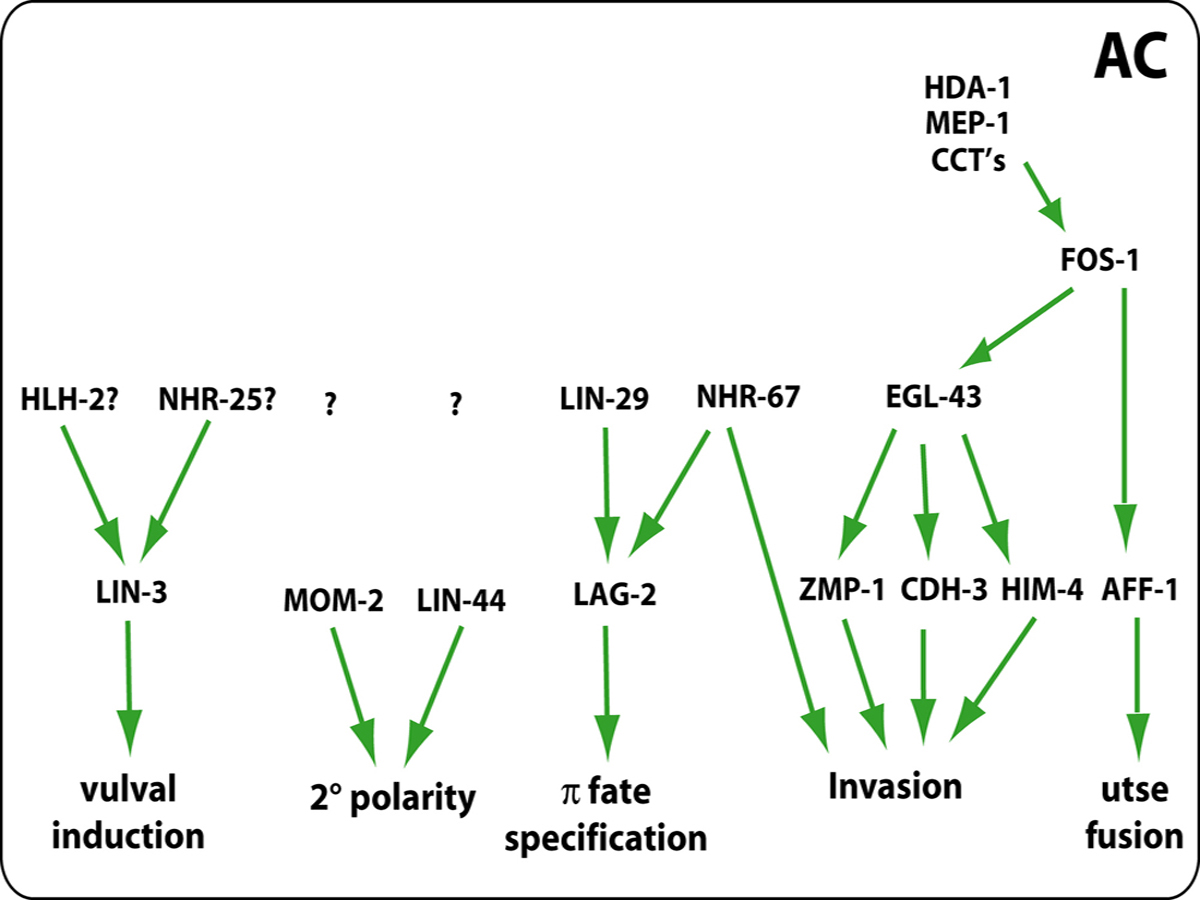
The regulatory circuitry controlling the anchor cell's many functions has started to be elucidated (Figure 8). Anchor cell expression of LIN-3, which induces the vulva, requires HLH-2 and NHR-25 or a related protein (Hwang and Sternberg, 2004). Anchor cell expression of LAG-2, which induces uterine π cells, requires LIN-29 and NHR-67 (Newman et al., 2000; Verghese et al., 2011). However, the regulation of anchor cell expression of the wnt proteins MOM-2 and LIN-44, which orient the polarity of the P5.p and P7.p lineages toward the anchor cell, is unknown.
|
Figure 8. Signaling network in the AC. The AC coordinates multiple processes in the egg-laying system by activating expression of several genes during L3 and L4 stages. In addition to producing LIN-3/EGF ligand that activates Ras signaling in VPCs, the AC is also the source of two Wnt ligands, MOM-2 and LIN-44, that control 2° lineage polarity. The invasion of AC into vulval epithelium is controlled by nhr-67, fos-1, and egl-43, and their downstream targets. In the case of utse fusion fos-1 controls aff-1 function. The upstream regulators of fos-1 include genes such as hda-1, mep-1 and cct complex.
Anchor cell invasion requires function of fos-1a (fos proto-oncogene family) (Sherwood et al., 2005), the long isoform of egl-43 (Evi1 proto-oncogene family) (Hwang et al., 2007; Rimann and Hajnal, 2007) and nhr-67 (NHR family) (Verghese et al., 2011). Histone deactylase HDA-1, zinc finger protein MEP-1 and components of the cct chaperonin complex function to control expression of fos-1a as well as efficient assembly of the invasive membrane (Matus et al., 2010).
FOS-1a is necessary for egl-43 expression but not vice versa (Hwang et al., 2007), and a FOS-1 responsive element in the egl-43 promoter contains a non-canonical binding site for FOS-1, suggesting direct regulation of egl-43 by FOS-1a (Rimann and Hajnal, 2007). Both FOS-1a and EGL-43 are required for expression of target genes zmp-1, cdh-3 and him-4 (Hwang et al., 2007; Rimann and Hajnal, 2007; Sherwood et al., 2005). The current model for the transcriptional network posits that EGL-43 regulation of these targets is direct and that FOS-1 may regulate them via EGL-43 and perhaps directly as well (Figure 8).
Considerable progress has been made in the last decade towards identifying cell type-specific-transcriptional networks, cell polaritzation mechanisms, and morphogenetic changes that form the vulva and the vulval-uterine connection. As a result we have learned that the polarity of the 2° lineage vulval cells is controlled by a β-Catenin-TCF/LEF-mediated Wnt signaling pathway. The vulval toroids are formed by coordinated morphogenetic events and selective cell fusion. We also have expanded our understanding of the role of the AC in the formation of vulval-uterine connection. In spite of this our mechanistic insights of these events is far from complete. For example, we know very little about the downstream targets of transcriptional networks and how they confer unique cell identities, how cell divisions and cell fates are coupled and, how the timing of different cellular events in the vulva and uterus are coordinated. Likewise, we have rudimentary knowledge of the mechanisms regulating morphogenetic processes such as cell adhesion and cell migration. We expect that in the near future the availability of various genetics and genomics tools will help find answers to some of these questions thereby providing a more coherent picture of cellular and molecular processes in the formation of the egg-laying system.
We thank members of our laboratories for discussion and feedback. BPG is supported by funds from NSERC, CIHR and Ministry of Research and Innovation (through ERA program). WHR is supported by R01 GM086786 from the NIH and IOS-071867 from the NSF. PWS is an investigator of the Howard Hughes Medical Institute.
Alkema, M. J., Hunter-Ensor, M., Ringstad, N., and Horvitz, H. R. (2005). Tyramine Functions independently of octopamine in the Caenorhabditis elegans nervous system. Neuron 46, 247-260. Abstract Article
Antoshechkin, I., and Han, M. (2002). The C. elegans evl-20 gene is a homolog of the small GTPase ARL2 and regulates cytoskeleton dynamics during cytokinesis and morphogenesis. Dev. Cell 2, 579-591. Abstract Article
Berninsone, P., Hwang, H. Y., Zemtseva, I., Horvitz, H. R., and Hirschberg, C. B. (2001). SQV-7, a protein involved in Caenorhabditis elegans epithelial invagination and early embryogenesis, transports UDP-glucuronic acid, UDP-N- acetylgalactosamine, and UDP-galactose. Proc. Natl. Acad. Sci. R. S. A. 98, 3738-3743. Abstract Article
Berset, T., Hoier, E. F., Battu, G., Canevascini, S., and Hajnal, A. (2001). Notch inhibition of RAS signaling through MAP kinase phosphatase LIP-1 during C. elegans vulval development. Science 291, 1055-1058. Abstract Article
Bettinger, J. C., Euling, S., and Rougvie, A. E. (1997). The terminal differentiation factor LIN-29 is required for proper vulval morphogenesis and egg laying in Caenorhabditis elegans. Development 124, 4333-4342. Abstract
Broday, L., Kolotuev, I., Didier, C., Bhoumik, A., Gupta, B. P., Sternberg, P. W., Podbilewicz, B., and Ronai, Z. (2004). The small ubiquitin-like modifier (SUMO) is required for gonadal and uterine-vulval morphogenesis in Caenorhabditis elegans. Genes Dev. 18, 2380-2391. Abstract Article
Burdine, R. D., Branda, C. S., and Stern, M. J. (1998). EGL-17(FGF) expression coordinates the attraction of the migrating sex myoblasts with vulval induction in C. elegans. Development 125, 1083-1093. Abstract
Chamberlin, H. M., Palmer, R. E., Newman, A. P., Sternberg, P. W., Baillie, D. L., and Thomas, J. H. (1997). The PAX gene egl-38 mediates developmental patterning in Caenorhabditis elegans. Development 124, 3919-3928. Abstract
Chang, C., Newman, A. P., and Sternberg, P. W. (1999). Reciprocal EGF signaling back to the uterus from the induced C. elegans vulva coordinates morphogenesis of epithelia. Curr. Biol. 9, 237-246. Abstract Article
Choi, J., Richards, K. L., Cinar, H. N., and Newman, A. P. (2006). N-ethylmaleimide sensitive factor is required for fusion of the C. elegans uterine anchor cell. Dev. Biol. 297, 87-102. Abstract Article
Cinar, H. N., Richards, K. L., Oommen, K. S., and Newman, A. P. (2003). The EGL-13 SOX domain transcription factor affects the uterine π cell lineages in Caenorhabditis elegans. Genetics 165, 1623-1628. Abstract
Colavita, A., and Tessier-Lavigne, M. (2003). A neurexin-related protein, BAM-2, terminates axonal branches in C. elegans. Science 302, 293-296. Abstract Article
Cui, M., and Han, M. (2003). Cis regulatory requirements for vulval cell-specific expression of the Caenorhabditis elegans fibroblast growth factor gene egl-17. Dev. Biol. 257, 104-116. Abstract Article
Dalpe, G., Brown, L., and Culotti, J. G. (2005). Vulva morphogenesis involves attraction of plexin 1-expressing primordial vulva cells to semaphorin 1a sequentially expressed at the vulva midline. Development 132, 1387-1400. Abstract Article
Deshpande, R., Inoue, T., Priess, J. R., and Hill, R. J. (2005). lin-17/Frizzled and lin-18 regulate POP-1/TCF-1 localization and cell type specification during C. elegans vulval development. Dev. Biol. 278, 118-129. Abstract Article
Eisenmann, D. M. (2005). Wnt signaling. WormBook ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.7.1, http://www.wormbook.org.
Eisenmann, D. M., and Kim, S. K. (2000). Protruding vulva mutants identify novel loci and Wnt signaling factors that function during Caenorhabditis elegans vulva development. Genetics 156, 1097-1116. Abstract
Estes, K. A., and Hanna-Rose, W. (2009). The anchor cell initiates dorsal lumen formation during C. elegans vulval tubulogenesis. Dev. Biol. 328, 297-304. Abstract Article
Estes, K. A., Kalamegham, R., and Hanna-Rose, W. (2007). Membrane localization of the NlpC/P60 family protein EGL-26 correlates with regulation of vulval cell morphogenesis in Caenorhabditis elegans. Dev. Biol. 308, 196-205. Abstract Article
Ferguson, E. L., Sternberg, P. W., and Horvitz, H. R. (1987). A genetic pathway for the specification of the vulval cell lineages of Caenorhabditis elegans. Nature 326, 259-267. Abstract Article
Fernandes, J. S., and Sternberg, P. W. (2007). The tailless ortholog nhr-67 regulates patterning of gene expression and morphogenesis in the C. elegans vulva. PLoS Genet. 3, e69. Abstract Article
Freyd, G. (1991). Molecular analysis of the Caenorhabditis elegans cell lineage gene lin-11, Ph.D., Massachusetts Institute of Technology, Boston, MA. Article
Green, J. L., Inoue, T., and Sternberg, P. W. (2008). Opposing Wnt pathways orient cell polarity during organogenesis. Cell 134, 646-656. Abstract Article
Greenwald, I. (2005). LIN-12/Notch signaling in C. elegans. WormBook ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.10.1, http://www.wormbook.org.
Gupta, B. P., and Sternberg, P. W. (2002). Tissue-specific regulation of the LIM homeobox gene lin-11 during development of the Caenorhabditis elegans egg-laying system. Dev. Biol. 247, 102-115. Abstract Article
Gupta, B. P., Wang, M., and Sternberg, P. W. (2003). The C. elegans LIM homeobox gene lin-11 specifies multiple cell fates during vulval development. Development 130, 2589-2601. Abstract Article
Hagedorn, E. J., Yashiro, H., Ziel, J. W., Ihara, S., Wang, Z., and Sherwood, D. R. (2009). Integrin acts upstream of netrin signaling to regulate formation of the anchor cell's invasive membrane in C. elegans. Dev. Cell 17, 187-198. Abstract Article
Hanna-Rose, W., and Han, M. (1999). COG-2, a Sox domain protein necessary for establishing a functional vulval-uterine connection in Caenorhabditis elegans. Development 126, 169-179. Abstract
Hanna-Rose, W., and Han, M. (2002). The Caenorhabditis elegans EGL-26 protein mediates vulval cell morphogenesis. Dev. Biol. 241, 247-258. Abstract Article
Hao, L., Johnsen, R., Lauter, G., Baillie, D., and Burglin, T. R. (2006). Comprehensive analysis of gene expression patterns of hedgehog-related genes. BMC Genomics 7, 280. Abstract Article
Herman, T., Hartwieg, E., and Horvitz, H. R. (1999). sqv mutants of Caenorhabditis elegans are defective in vulval epithelial invagination. Proc. Natl. Acad. Sci. R. S. A. 96, 968-973. Abstract Article
Herman, T., and Horvitz, H. R. (1999). Three proteins involved in Caenorhabditis elegans vulval invagination are similar to components of a glycosylation pathway. Proc. Natl. Acad. Sci. R. S. A. 96, 974-979. Abstract Article
Huang, L., and Hanna-Rose, W. (2006). EGF signaling overcomes a uterine cell death associated with temporal mis-coordination of organogenesis within the C. elegans egg-laying apparatus. Dev. Biol. 300, 599-611. Abstract Article
Hurd, D. D., and Kemphues, K. J. (2003). PAR-1 is required for morphogenesis of the Caenorhabditis elegans vulva. Dev. Biol. 253, 54-65. Abstract Article
Hwang, B. J., Meruelo, A. D., and Sternberg, P. W. (2007). C. elegans EVI1 proto-oncogene, EGL-43, is necessary for Notch-mediated cell fate specification and regulates cell invasion. Development 134, 669-679. Abstract Article
Hwang, B. J., and Sternberg, P. W. (2004). A cell-specific enhancer that specifies lin-3 expression in the C. elegans anchor cell for vulval development. Development 131, 143-151. Abstract Article
Hwang, H. Y., and Horvitz, H. R. (2002). The Caenorhabditis elegans vulval morphogenesis gene sqv-4 encodes a UDP-glucose dehydrogenase that is temporally and spatially regulated. Proc. Natl. Acad. Sci. U. S. A. 99, 14224-14229. Abstract Article
Hwang, H. Y., and Horvitz, H. R. (2002). The SQV-1 UDP-glucuronic acid decarboxylase and the SQV-7 nucleotide-sugar transporter may act in the Golgi apparatus to affect Caenorhabditis elegans vulval morphogenesis and embryonic development. Proc. Natl. Acad. Sci. U. S. A. 99, 14218-14223. Abstract Article
Hwang, H. Y., Olson, S. K., Brown, J. R., Esko, J. D., and Horvitz, H. R. (2003). The Caenorhabditis elegans genes sqv-2 and sqv-6, which are required for vulval morphogenesis, encode glycosaminoglycan galactosyltransferase II and xylosyltransferase. J. Biol. Chem. 278, 11735-11738. Abstract Article
Hwang, H. Y., Olson, S. K., Esko, J. D., and Horvitz, H. R. (2003). Caenorhabditis elegans early embryogenesis and vulval morphogenesis require chondroitin biosynthesis. Nature 423, 439-443. Abstract Article
Inoue, T., Oz, H. S., Wiland, D., Gharib, S., Deshpande, R., Hill, R. J., Katz, W. S., and Sternberg, P. W. (2004). C. elegans LIN-18 is a Ryk ortholog and functions in parallel to LIN-17/Frizzled in Wnt signaling. Cell 118, 795-806. Abstract Article
Inoue, T., Sherwood, D. R., Aspock, G., Butler, J. A., Gupta, B. P., Kirouac, M., Wang, M., Lee, P. Y., Kramer, J. M., Hope, I., et al. (2002). Gene expression markers for Caenorhabditis elegans vulval cells. Mech. Dev. 119 Suppl 1, S203-209. Abstract Article
Inoue, T., and Sternberg, P. W. (2010). C. elegans BED domain transcription factor BED-3 controls lineage-specific cell proliferation during organogenesis. Dev. Biol. 338, 226-236. Abstract Article
Inoue, T., Wang, M., Ririe, T. O., Fernandes, J. S., and Sternberg, P. W. (2005). Transcriptional network underlying Caenorhabditis elegans vulval development. Proc. Natl. Acad. Sci. R. S. A. 102, 4972-4977. Abstract Article
Johnson, R. W., Liu, L. Y., Hanna-Rose, W., and Chamberlin, H. M. (2009). The Caenorhabditis elegans heterochronic gene lin-14 coordinates temporal progression and maturation in the egg-laying system. Dev. Dyn. 238, 394-404. Abstract Article
Jose, A. M., Bany, I. A., Chase, D. L., and Koelle, M. R. (2007). A specific subset of transient receptor potential vanilloid-type channel subunits in Caenorhabditis elegans endocrine cells function as mixed heteromers to promote neurotransmitter release. Genetics 175, 93-105. Abstract Article
Katz, W. S., Lesa, G. M., Yannoukakos, D., Clandinin, T. R., Schlessinger, J., and Sternberg, P. W. (1996). A point mutation in the extracellular domain activates LET-23, the Caenorhabditis elegans epidermal growth factor receptor homolog. Mol. Cell. Biol. 16, 529-537. Abstract
Kim, K., and Li, C. (2004). Expression and regulation of an FMRFamide-related neuropeptide gene family in Caenorhabditis elegans. J. Comp. Neurol. 475, 540-550. Abstract Article
Kimble, J., and Hirsh, D. (1979). The postembryonic cell lineages of the hermaphrodite and male gonads in Caenorhabditis elegans. Dev. Biol. 70, 396-417. Abstract Article
Kirouac, M., and Sternberg, P. W. (2003). cis-Regulatory control of three cell fate-specific genes in vulval organogenesis of Caenorhabditis elegans and C. briggsae. Dev. Biol. 257, 85-103. Abstract Article
Kishore, R. S., and Sundaram, M. V. (2002). ced-10 Rac and mig-2 function redundantly and act with unc-73 trio to control the orientation of vulval cell divisions and migrations in Caenorhabditis elegans. Dev. Biol. 241, 339-348. Abstract Article
Klerkx, E. P., Alarcon, P., Waters, K., Reinke, V., Sternberg, P. W., and Askjaer, P. (2009). Protein kinase VRK-1 regulates cell invasion and EGL-17/FGF signaling in Caenorhabditis elegans. Dev. Biol. 335, 12-21. Abstract Article
Kontani, K., Moskowitz, I. P., and Rothman, J. H. (2005). Repression of cell-cell fusion by components of the C. elegans vacuolar ATPase complex. Dev. Cell 8, 787-794. Abstract Article
Li, C., and Chalfie, M. (1990). Organogenesis in C. elegans: positioning of neurons and muscles in the egg-laying system. Neuron 4, 681-695. Abstract Article
Marri, S., and Gupta, B. P. (2009). Dissection of lin-11 enhancer regions in Caenorhabditis elegans and other nematodes. Dev. Biol. 325, 402-411. Abstract Article
Matus, D. Q., Li, X. Y., Durbin, S., Agarwal, D., Chi, Q., Weiss, S. J., and Sherwood, D. R. (2010). In vivo identification of regulators of cell invasion across basement membranes. Sci. Signaling 3, ra35. Abstract Article
Mohler, W. A., Shemer, G., del Campo, J. J., Valansi, C., Opoku-Serebuoh, E., Scranton, V., Assaf, N., White, J. G., and Podbilewicz, B. (2002). The type I membrane protein EFF-1 is essential for developmental cell fusion. Dev. Cell 2, 355-362. Abstract Article
Newman, A. P., Acton, G. Z., Hartwieg, E., Horvitz, H. R., and Sternberg, P. W. (1999). The lin-11 LIM domain transcription factor is necessary for morphogenesis of C. elegans uterine cells. Development 126, 5319-5326. Abstract
Newman, A. P., Inoue, T., Wang, M., and Sternberg, P. W. (2000). The Caenorhabditis elegans heterochronic gene lin-29 coordinates the vulval-uterine-epidermal connections. Curr. Biol. 10, 1479-1488. Abstract Article
Newman, A. P., White, J. G., and Sternberg, P. W. (1995). The Caenorhabditis elegans lin-12 gene mediates induction of ventral uterine specialization by the anchor cell. Development 121, 263-271. Abstract
Newman, A. P., White, J. G., and Sternberg, P. W. (1996). Morphogenesis of the C. elegans hermaphrodite uterus. Development 122, 3617-3626. Abstract
Oommen, K. S., and Newman, A. P. (2007). Co-regulation by Notch and Fos is required for cell fate specification of intermediate precursors during C. elegans uterine development. Development 134, 3999-4009. Abstract Article
Palmer, R. E., Inoue, T., Sherwood, D. R., Jiang, L. I., and Sternberg, P. W. (2002). Caenorhabditis elegans cog-1 locus encodes GTX/Nkx6.1 homeodomain proteins and regulates multiple aspects of reproductive system development. Dev. Biol. 252, 202-213. Abstract Article
Pellegrino, M. W., Farooqui, S., Frohli, E., Rehrauer, H., Kaeser-Pebernard, S., Muller, F., Gasser, R. B., and Hajnal, A. (2011). LIN-39 and the EGFR/RAS/MAPK pathway regulate C. elegans vulval morphogenesis via the VAB-23 zinc finger protein. Development 138, 4649-4660. Abstract Article
Perens, E. A., and Shaham, S. (2005). C. elegans daf-6 encodes a patched-related protein required for lumen formation. Dev. Cell 8, 893-906. Abstract Article
Pettitt, J., Wood, W. B., and Plasterk, R. H. (1996). cdh-3, a gene encoding a member of the cadherin superfamily, functions in epithelial cell morphogenesis in Caenorhabditis elegans. Development 122, 4149-4157. Abstract
Rajakumar, V., and Chamberlin, H. M. (2007). The Pax2/5/8 gene egl-38 coordinates organogenesis of the C. elegans egg-laying system. Dev. Biol. 301, 240-253. Abstract Article
Reece-Hoyes, J. S., Shingles, J., Dupuy, D., Grove, C. A., Walhout, A. J., Vidal, M., and Hope, I. A. (2007). Insight into transcription factor gene duplication from Caenorhabditis elegans Promoterome-driven expression patterns. BMC Genomics 8, 27. Abstract Article
Rimann, I., and Hajnal, A. (2007). Regulation of anchor cell invasion and uterine cell fates by the egl-43 Evi-1 proto-oncogene in Caenorhabditis elegans. Dev. Biol. 308, 187-195. Abstract Article
Ririe, T. O., Fernandes, J. S., and Sternberg, P. W. (2008). The Caenorhabditis elegans vulva: a post-embryonic gene regulatory network controlling organogenesis. Proc. Natl. Acad. Sci. R. S. A. 105, 20095-20099. Abstract Article
Sapir, A., Choi, J., Leikina, E., Avinoam, O., Valansi, C., Chernomordik, L. V., Newman, A. P., and Podbilewicz, B. (2007). AFF-1, a FOS-1-regulated fusogen, mediates fusion of the anchor cell in C. elegans. Dev. Cell 12, 683-698. Abstract Article
Sawa, H., Lobel, L., and Horvitz, H. R. (1996). The Caenorhabditis elegans gene lin-17, which is required for certain asymmetric cell divisions, encodes a putative seven-transmembrane protein similar to the Drosophila frizzled protein. Genes Dev. 10, 2189-2197. Abstract Article
Seetharaman, A., Cumbo, P., Bojanala, N., and Gupta, B. P. (2010). Conserved mechanism of Wnt signaling function in the specification of vulval precursor fates in C. elegans and C. briggsae. Dev. Biol. 346, 128-139). Abstract Article
Sharma-Kishore, R., White, J. G., Southgate, E., and Podbilewicz, B. (1999). Formation of the vulva in Caenorhabditis elegans: a paradigm for organogenesis. Development 126, 691-699. Abstract
Shen, K., and Bargmann, C. I. (2003). The immunoglobulin superfamily protein SYG-1 determines the location of specific synapses in C. elegans. Cell 112, 619-630. Abstract Article
Shen, K., Fetter, R. D., and Bargmann, C. I. (2004). Synaptic specificity is generated by the synaptic guidepost protein SYG-2 and its receptor, SYG-1. Cell 116, 869-881. Abstract Article
Sherwood, D. R., Butler, J. A., Kramer, J. M., and Sternberg, P. W. (2005). FOS-1 promotes basement-membrane removal during anchor-cell invasion in C. elegans. Cell 121, 951-962. Abstract Article
Sherwood, D. R., and Sternberg, P. W. (2003). Anchor cell invasion into the vulval epithelium in C. elegans. Dev. Cell 5, 21-31. Abstract Article
Sternberg, P. W. (2005). Vulval development. WormBook ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.6.1, http://www.wormbook.org. Abstract Article
Sternberg, P. W., and Horvitz, H. R. (1988). lin-17 mutations of Caenorhabditis elegans disrupt certain asymmetric cell divisions. Dev. Biol. 130, 67-73. Abstract Article
Stringham, E., Pujol, N., Vandekerckhove, J., and Bogaert, T. (2002). unc-53 controls longitudinal migration in C. elegans. Development 129, 3367-3379. Abstract
Thomas, J. H., Stern, M. J., and Horvitz, H. R. (1990). Cell interactions coordinate the development of the C. elegans egg-laying system. Cell 62, 1041-1052. Abstract Article
Ulm, E. A., Sleiman, S. F., and Chamberlin, H. M. (2011). Developmental functions for the Caenorhabditis elegans Sp protein SPTF-3. Mech. Dev. 128, 428-441. Abstract Article
Verghese, E., Schocken, J., Jacob, S., Wimer, A. M., Royce, R., Nesmith, J. E., Baer, G. M., Clever, S., McCain, E., Lakowski, B., and Wightman, B. (2011). The tailless ortholog nhr-67 functions in the development of the C. elegans ventral uterus. Dev. Biol. 356, 516-528. Abstract Article
Wagmaister, J. A., Miley, G. R., Morris, C. A., Gleason, J. E., Miller, L. M., Kornfeld, K., and Eisenmann, D. M. (2006). Identification of cis-regulatory elements from the C. elegans Hox gene lin-39 required for embryonic expression and for regulation by the transcription factors LIN-1, LIN-31 and LIN-39. Dev. Biol. 297, 550-565. Abstract Article
Wang, M., and Sternberg, P. W. (2000). Patterning of the C. elegans primary vulval lineage by RAS and Wnt pathways. Development 127, 5047-5058. Abstract
Zand, T. P., Reiner, D. J., and Der, C. J. (2011). Ras effector switching promotes divergent cell fates in C. elegans vulval patterning. Dev. Cell 20, 84-96. Abstract Article
*Edited by Barbara Meyer. Last revised June 8, 2012. Published November 30, 2012. This chapter should be cited as: Gupta, B. P. et al. Morphogenesis of the vulva and the vulval-uterine connection (November 30, 2012), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/wormbook.1.152.1, http://www.wormbook.org.
Copyright: © 2012 Gupta et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
§To whom correspondence should be address. E-mail: guptab@mcmaster.ca
 All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.
All WormBook content, except where otherwise noted, is licensed under a Creative Commons Attribution License.